Лекция по теме "Пластический обмен"
Лекция 6. Пластический обмен в клетке.Пластический обмен – к пластическому обмену относится синтез нуклеиновых кислот, биосинтез белков, фотосинтез, жиров и углеводов. В клетке есть два типа нуклеиновых кислот: ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). ДНК и РНК состоят из нуклеотидов. Каждый нуклеотид состоит из: 1. азотистого основания (аденина, гуанина, цитозина, тимина или урацила), 2. углевода (дезоксирибозы или рибозы) 3. остатка фосфорной кислоты (Н2РО4). Цепочки нуклеотидов связаны водородными связями. ДНК – это основной носитель информации о структуре белков в клетке. ДНК состоит из двух цепочек нуклеотидов, закрученных одна вокруг другой в спираль. ДНК находится в ядре, в митохондриях, в пластидах. Двойная спираль ДНК построена по принципу комплиментарности (комплиментарность – взаимное дополнение). Молекулы ДНК обладают свойством – способностью к удвоению. Принцип комплиментарности лежит и в основе удвоения молекул ДНК. С помощью специальных ферментов водородные связи, скрепляющие нити ДНК, разрываются, и к каждому нуклеотиду каждой из этих нитей последовательно пристраиваются комплиментарные нуклеотиды. Разошедшиеся нити исходной (материнской) молекулы ДНК являются матричными. Они задают порядок расположения нуклеотидов во вновь синтезируемой цепи. В результате действия сложного набора ферментов происходит соединение нуклеотидов друг с другом. При этом образуются новые нити ДНК, комплиментарные каждой из разошедшихся цепей. Таким образом, в результате удвоения создаются две двойные спирали ДНК (дочерние молекулы), каждая из которых имеет одну нить, полученную от материнской молекулы, и одну нить, синтезированную вновь. Этапы удвоения ДНК: 1. разрыв водородных связей, скрепляющих нити ДНК; расхождение нитей ДНК, 2. подбор к материнским нитям ДНК комплементарных нуклеотидов, 3. синтез дочерних ДНК, Процесс самоудвоения молекулы ДНК называют репликацией. При этом не расходуется энергия АТФ. Дочерние молекулы ДНК имеют одинаковые гены, какие были у материнской. Каждая клетка многоклеточного организма возникает из одной зародышевой клетки в результате многократных делений. Следовательно, все клетки организма имеют одинаковый набор генов. Случайно возникшая ошибка в гене зародышевой клетки будет воспроизведена в генах миллионов её потомков.
Образование информационной РНК по матрице ДНК К рибосомам, местам синтеза белков, из ядра поступает несущей информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). Это одноцепочная молекула, комплиментарная одной нити молекулы ДНК. Специальный фермент – полимераза, двигаясь по ДНК, подбирает по принципу комплиментарности нуклеотиды и соединяет их в единые цепочки. Этот процесс называется транскрипцией. Если в нити ДНК стоит Тимин, то полимераза включает в цепь иРНК Аденин, если стоит Гуанин – включает Цитозин, если Аденин – Урацил (в состав РНК не входит Тимин). иРНК – копия не всей молекулы ДНК, а только части её, одного гена или группы рядом стоящих генов, несущих информацию о структуре белков, необходимых для выполнения одной функции. У прокариот такая группа генов называется опероном. Вначале каждой группы генов находится своего рода посадочная площадка для полимеразы, называемая промотором. Это специфичная последовательность нуклеотидов ДНК, которую фермент “узнаёт” благодаря химическому сродству. Только присоединившись к промотору, полимераза способна начать синтез иРНК. В конце группы генов фермент встречает сигнал (в виде определённой последовательности нуклеотидов), означающий конец переписывания. Готовая иРНК отходит от ДНК, покидает ядро и направляется к месту синтеза белков – рибосоме, расположенной в цитоплазме клетки. В клетке генетическая информация передаётся благодаря транскрипции от ДНК к белку: ДНК → иРНК→ белок.
Синтез белков в клетке.Важнейшим процессом ассимиляции является синтез белков. Так как они выполняют в организме целый ряд функций, являются основой уникальности каждого вида. Ген — участок молекулы ДНК, характеризующийся определенной последовательностью нуклеотидов, в котором закодирована информация о первичной структуре полипептидной цепи (последовательности аминокислот в конкретном белке) или нуклеотидов в РНК. В одной молекуле ДНК содержится от сотен до десятков тысяч генов. Генетический код — это единая система записи наследственной информации в молекулах нуклеиновых кислот ДНК и и-РНК и виде последовательности нуклеотидов. Свойства генетического кода: 1. триплетность: каждая аминокислота кодируется определенным триплетом (или кодоном) — сочетанием трех последовательно расположенных нуклеотидов; 2. множественность (или избыточность): одна и та же аминокислота может кодироваться несколькими различными триплетами (от 2 до 6); 3. однозначность: каждый триплет кодирует только одну аминокислоту; 4. неперекрываемость: один нуклеотид не может входить в состав соседних триплетов; 5. непрерывность: гены в цепи нуклеотидов имеют строго фиксированные стартовые (или инициирующие) кодоны и терминирующие кодоны, сигнализирующие об окончании синтеза полипептидной цепи; внутри последовательности нуклеотидов гена «знаки препинания» отсутствуют; 6. универсальность: одинаковые триплеты кодируют одну и ту же аминокислоту у всех живых организмов. Матричный синтез — синтез молекул сложных органических веществ (белка, РНК, ДНК) из более простых на основе генетической информации, закодированной на матрице. Матрица — это готовая структура (молекула ДНК или и-РНК), содержащая закодированную генетическую информацию, в соответствии с которой осуществляется синтез новой структуры. Кодон — три рядом расположенных нуклеотида в молекулах ДНК или и-РНК, кодирующие одну аминокислоту. Транскрипция — процесс «считывания» генетической информации с молекулы ДНК и копирование ее на молекулу и-РНК.
Механизм транскрипции: фермент РНК-полимераза раскручивает двойную спираль молекулы ДНК на участке, соответствующем определенному гену, и обнажает одну из цепей спирали. Двигаясь вдоль этой цепи и встретив инициирующий кодон, РНК-полимераза начинает подбирать в кариоплазме нуклеотиды, комплементарные нуклеотидам гена ДНК, и соединяет их в цепочку и-РНК (молекулы информационной РНК). Процесс завершается после того как РНК-полимераза встретит в цепочке нуклеотидов ДНК терминирующий кодон. Таким образом, в результате транскрипции последовательность нуклеотидов, расположенных на участке от инициирующего до терминирующего кодона, «переписывается» в последовательность нуклеотидов и-РНК.
Строение молекулы т-РНК.
Благодаря определенному расположению комплементарных нуклеотидов и образованию между некоторыми из них водородных связей молекула т-РНК напоминает по форме лист клевера. На ее верхушке расположен антикодон -триплет свободных нуклеотидов, ответственный за узнавание соответствующего (комплементарного ему) кодона молекулы и-РНК. Основание молекулы т-РНК является акцептором, т.е. служит местом прикрепления именно той и только той аминокислоты, которой соответствует антикодон данной молекулы т-РНК. Механизм рекогниции (распознавания) для того чтобы молекула т-РНК могла присоединить к своему акцепторному концу аминокислоту, необходимо, чтобы аминокислота была активирована, т.е. имела определенную избыточную энергию. Активация аминокислот происходит в цитоплазме с помощью специального фермента, который расщепляет молекулы АТФ и передают выделившуюся при этом энергию молекулам аминокислот. Молекула т-РНК выбирает из цитоплазмы соответствующую ее антикодону активированную аминокислоту и переносит ее в рибосому. Одна молекула т-РНК может транспортировать только одну аминокислоту. Трансляция — это второй этап синтеза белка, выполняемый рибосомами по принципу комплементарное кодона и-РНК и антикодона т-РНК. В процессе трансляции осуществляется расшифровка генетической информации, переносимой молекулами и-РНК, и «перевод» ее с нуклеотидного кода на аминокислотный. Механизм трансляции. Для трансляции необходимо, чтобы цепь и-РНК оказалась в канале, образующемся между меньшей и большей субъединицами рибосомы. В процессе трансляции эта цепь движется по каналу, так что в нем в каждый момент времени находится всего два кодона молекулы и-РНК. Трансляция начинается с инициации, когда через канал рибосомы пройдет стартовая аминокислота. В большую субъединицу рибосомы непрерывно поступают комплексы т-РНК + аминокислота, которые сменяют друг друга, причем в любой момент времени там находятся два комплекса, расположенные рядом. Если антикодон т-РНК оказывается комплементарным кодону и-РНК, то комплекс т-РНК + аминокислота временно присоединяется к цепочке и-РНК. Ко второму кодону и-РНК присоединяется второй комплекс т-РНК + аминокислота. С помощью ферментов между аминокислотами этих комплексов устанавливается пептидная связь и одновременно разрушаются связи между первой аминокислотой и т-РНК и между первой т-РНК и цепочкой и-РНК. т-РНК уходит из рибосомы за следующей аминокислотой, а цепочка и-РНК сдвигается на один триплет, и процесс повторяется.
В результате каждого такого шага молекула будущего белка увеличивается на одну аминокислоту в строгом соответствии с порядком, указанным молекулой и-РНК. Синтез полипептидной белковой цепи завершается тогда, когда в рибосому попадут терминирующие кодоны и-РНК. После этого полипептидная белковая молекула отделяется от рибосомы и поступает в канальцы ЭПС, где приобретает свойственную ей пространственную структуру. Одна молекула и-РНК позволяет считывать с себя информацию сразу нескольким рибосомам. Полисома — это комплекс, состоящий из и-РНК и нескольких (от 5-6 до нескольких десятков) рибосом. Полисомы позволяют одновременно осуществлять синтез нескольких полипептидных цепей. Синтез белковых молекул происходит непрерывно; за 1 мин образуется 50-60 тыс. пептидных связей. Одна молекула белка синтезируется за 3-4 с.
После завершения синтеза белковая молекула отделяется от рибосомы и приобретает свойственную ей (вторичную, третичную или четвертичную) структуру. Скорость биосинтеза зависит от активности ферментов, катализирующих процессы транскрипции и трансляции, от температуры, концентрации водородных ионов, наличия АТФ и свободных аминокислот и др. В клетках есть специальные механизмы, регулирующие активность генов, благодаря чему в каждый данный момент синтезируются только те белки, которые ей необходимы.
|
 Каждый триплет нуклеотидов и-РНК является кодоном, по которому в процессе сборки молекулы белка будет подбираться соответствующая аминокислота.
Каждый триплет нуклеотидов и-РНК является кодоном, по которому в процессе сборки молекулы белка будет подбираться соответствующая аминокислота.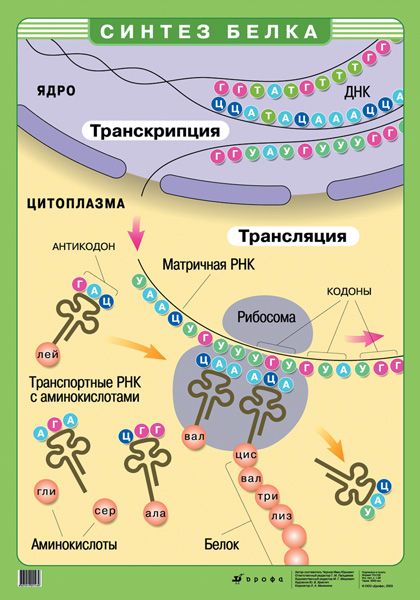

Фотосинтез — это процесс синтеза органических веществ из молекул углекислого газа и воды, происходящий с использованием энергии света (обычно солнечной энергии) в зеленых растениях, некоторых бактериях и сопровождающийся выделением кислорода.
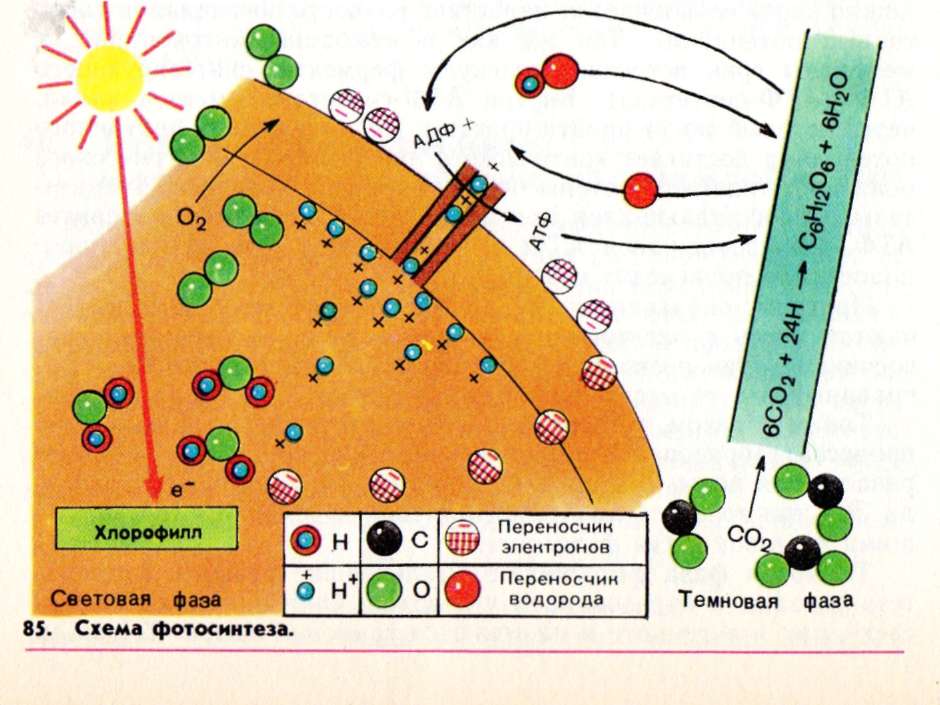
Фотосинтезирующими органоидами являются хлоропласты. Структурной и функциональной единицей хлоропластов являются тилакоиды – плоские мембранные мешочки, уложенные в стопки. На мембранах тилакоидов расположены комплексы с молекулами хлорофилла. Хлолрофилл способен улавливать кванты света. Коэффициент полезного действия фотосинтеза ~60%. Уравнение фотосинтеза:
6СO2 + 6Н2O + свет → С6Н12O6 + 6O2↑.
Стадии фотосинтеза: световая (осуществляется в тилакоидах гран) и темновая (осуществляется в строме хлоропластов).
Процессы световой фазы
Видимый свет частично поглощается хлорофиллом, в результате чего некоторые его молекулы возбуждаются и теряют электроны е—, превращаясь в положительно заряженные ионы. Одновременно под действием света происходит фотолиз (фоторазложение) воды с образованием ионов ОН— и Н+ : Н2O → ОН— + Н+ . Ионы Н— накапливаются преимущественно на внутренней стороне мембраны, заряжая ее положительно. Некоторые гидроксильные группы ОН— теряют электроны, восстанавливаясь до радикалов ОН, которые объединяются, образуя воду и свободный кислород, выделяемый в атмосферу:
ОН— → ОН + е—, 4OН → 2Н2О +O2↑.
Часть электронов, потерянных возбужденным хлорофиллом и гидроксилом, пройдя по электронно-транспортной цепи мембраны, накапливается преимущественно на ее внешней стороне, заряжая ее отрицательно. Оставшиеся электроны рекомбинируют с частью образовавшихся положительно заряженных ионов хлорофилла. В результате разделения заряженных частиц е— и Н+ между внешней и внутренней сторонами мембраны образуется электрическое поле. Когда оно достигает некоторой критической величины, ионы Н+ (протоны) устремляются по протонному каналу в ферменте АТФ-синтетаза, встроенному в мембрану тилакоида, к внешней поверхности мембраны. Достигнув ее, они соединяются с электронами, образуя атомарный водород: Н+е—→Н. При этом выделяется энергия, которая идет на синтез молекул АТФ. Образовавшиеся молекулы АТФ переходят в строму хлоропласта. Нейтральные атомы водорода Н соединяются с молекулами кофермента, кратко называемого НАДФ , образуя комплексы НАДФ*Н+ Н+, которые также переходят в строму.
Результаты процессов световой фазы: образование молекул АТФ, комплексов НАДФ*Н+ Н+ и свободного кислорода О2.
Темновая фаза
Реакции протекают во внутренней среде хлоропластов. Могут происходить как при наличии света, так и без него. Синтезируются органические вещества с использованием энергии, которая образовалась в световой фазе.
Процесс восстановления углекислого газа является циклическим и называется циклом Кальвина. Назван в честь американского исследователя М. Кальвина, который открыл этот циклический процесс.
Процессы темновой фазы происходят в строме хлоропласта, куда поступает АТФ, НАДФ*Н+ Н+ и СО2 (через устьица из воздуха). В присутствии ферментов молекулы СО2 присоединяются к молекулам присутствующего в строме сахара-пентозы рибупозодифосфата (С5). При этом образуется нестойкое шестиуглеродное соединение (С6), которое ферментативным путем распадается на две триозы (С3) — трехуглеродные молекулы фосфоглицериновой кислоты и фосфоглицеринового альдегида (которые для краткости обозначим ФГ). Превращения молекул ФГ происходят при участии продуктов световой фазы (АТФ и комплексов НАДФ*Н+ Н+). Каждая из молекул ФГ отнимает по одной фосфатной группе у молекулы АТФ, обогащаясь при этом энергией, а затем отщепляет атомы водорода от НАДФ*Н+Н+ , окисляя его до НАДФ. Дальнейшие превращения молекул ФГ осуществляются по одному из трех вариантов. Одна часть этих молекул объединяется, образуя углеводы (глюкозу) и воду; полученные углеводы затем могут полимеризоваться, образуя крахмал, целлюлозу и т.п. Другая часть участвует в синтезе аминокислот, карбоновых кислот, спиртов и др. Третья часть молекул ФГ участвует в цепочке реакций, в результате которых триозы превращаются в пятиуглеродные молекулы исходного вещества — рибулозодифосфата, тем самым замыкая цикл химических превращений — C3-цикл, или цикл Кальвина
Итоговое уравнение темновой фазы: 6СО2 + 24Н → С6Н12О6 +6Н2О

Значение фотосинтеза: он основной источник первичного органического вещества, единственный источник свободного кислорода на Земле и регулятор содержания СО2 в атмосфере; энергия, полученная от Солнца и запасенная в химических связях органических соединений, используется всеми гетеротрофными организмами.