ФИЗИОЛОГИЧЕСКИЕ РОЛИ ЧАСТНЫХ ОБРАЗОВАНИЙ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
ФИЗИОЛОГИЧЕСКИЕ РОЛИ ЧАСТНЫХ ОБРАЗОВАНИЙ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
План лекции
1. Частная физиология спинного мозга
2. Частная физиология продолговатого мозга и варолиева моста
3. Частная физиология среднего мозга
4. Частная физиология мозжечка
5. Частная физиология ретикулярной формации
6. Частная физиология промежуточного мозга
7. Частная физиология лимбической системы
8. Частная физиология подкорковых ядер
9. Частная физиология коры больших полушарий
10. .Периферический соматический отдел нервной системы
11. Вегетативный отдел нервной системы
12. Принцип деятельности вегетативного отдела нервной системы.
Центральная нервная система — это головной и спинной мозг. Головной мозг включает в себя задний мозг, или продолговатый мозг и варолиев мост, средний мозг, ретикулярную формацию, мозжечок, промежуточный мозг, лимбическую систему, подкорковые ядра, кору больших полушарий (рис. 1).
1.СПИННОЙ МОЗГ
Спинной мозг расположен в позвоночном канале и представляет собой цилиндрический тяж с дорсальными и вентральными корешками. Он переходит в ствол головного мозга. Спинной мозг — структурно-физиологическое образование ЦНС из нейронов. Тела нейронов формируют серое вещество спинного мозга (расположено в центре), а отростки нейронов — белое вещество (расположено по периферии). По физиологической роли различают три вида нейронов спинного мозга: промежуточные (интернейроны), моторные (эффекторные) и вегетативные (рис. 1). Нейроны спинного мозга формируют исполнительные отделы нервных центров рефлекторных дуг ряда рефлексов. Вентральные корешки называют двигательными, так как они содержат
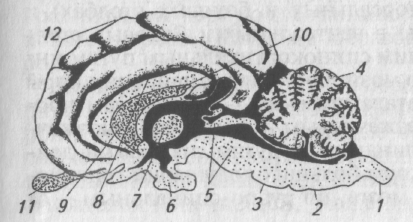
Рис. 1. Основные отделы центральной нервной системы:
1 — спинной мозг; 2— продолговатый мозг; 3 — варолиев мост; 4 — мозжечок; 5—средний мозг; 6— гипоталамус; 7— таламус, 8— лимбическая система; 9— полосатое тело; 10— гиппокамп; 11—обонятельная луковица; 12— кора больших полушарий

Рис. 2. Спинной мозг (поперечный разрез):
Показаны контакты между рецепторными, вставочными (промежуточными) и двигательными (моторными) нейронами в сером веществе спинного мозга отростки (аксоны) двигательных нейронов (сами тела нейронов расположены в сером веществе спинного мозга), иннервирующие скелетные мышцы; дорсальные корешки — чувствительными: состоят из отростков рецепторных нейронов (сами тела нейронов расположены в спинномозговых узлах).
Рефлекторная деятельность спинного мозга. Спинной мозг получает информацию с рецепторов кожи, мышц, туловища и конечностей, внутренних органов. Информация с рецепторов поступает к центрам спинного мозга. В спинном мозге находятся исполнительные отделы нервных центров, с участием которых осуществляется целый ряд наиболее простых и сложных рефлексов: 1) сгибания и разгибания конечностей; 2) потоотделения; 3) мочеиспускания; 4) дефекации; 5) молоковыведения, 6) эрекции полового члена; 7) эякуляции; 8) сердечно-сосудистых, дыхательных, пищевых, обмена веществ. Все рефлексы спинного мозга в естественных условиях осуществляются с участием головного мозга, включая кору больших полушарий.
Проводниковая деятельность спинного мозга. Она осуществляется за счет наличия в спинном мозге проводящих путей, которые образованы промежуточными нейронами. Проводящие пути структурно-функционально соединяют нейроны спинного мозга с нейронами других отделов ЦНС. Проводящие пути делят на восходящие (расположены в дорсальных и боковых столбах) и нисходящие (расположены в вентральных и боковых столбах) пути. По парным восходящим спинокортикальным путям информация с нейронов спинного мозга поступает к нейронам коры больших полушарий, по спиноталамическим — к нейронам промежуточного мозга, по спиномозжечковым — к нейронам мозжечка. По нисходящим кортикоспинальным путям программа действия передается от нейронов коры больших полушарий головного мозга к нейронам спинного мозга, по руброспинальным — от нейронов красного ядра среднего мозга, по вестибулоспинальным — от вестибулярных ядер продолговатого мозга, по ретикулоспинальным — от нейронов ретикулярной формации, по тектоспинальным — от нейронов бугров четверохолмия к нейронам спинного мозга. В итоге обеспечиваются полноценные приспособительные соматические и вегетативные реакции организма.
2. ПРОДОЛГОВАТЫЙ МОЗГ И ВАРОЛИЕВ МОСТ
Продолговатый мозг и варолиев мост — структурно-физиологическое образование ЦНС; образованы нейронами. Они, объединяясь, образуют ядра ряда черепно-мозговых нервов — тройничных, отводящих, лицевых, слуховых, языкоглоточных, блуждающих, добавочных, подъязычных и соответственно нервные центры, эфферентные звенья рефлекторных дуг ряда рефлексов. В черепномозговых ганглиях располагаются рецепторные нейроны, образующие афферентные звенья рефлекторных дуг ряда рефлексов. Продолговатому мозгу присущи два вида деятельности: рефлекторная и проводниковая. Ему характерна большая сложность выполняемых функций, чем спинному. Все реакции, осуществляемые продолговатым мозгом, более сложные.
Рефлекторная деятельность. Скопления нейронов продолговатого мозга образуют нервные центры, осуществляющие следующие жизненно важные рефлексы: дыхания, сердечно-сосудистый, пищевой, сосания, жевания, глотания, мигания, кашля, чихания, слезоотделения, рвоты, углеводного обмена, потоотделения, тонуса мышц (обеспечивающего естественную позу).
Проводниковая деятельность. Нейроны продолговатого мозга и варолиева моста связаны с нейронами спинного мозга и всех других отделов ЦНС посредством проводящих путей. От них идут ретикулоспинальный и вестибулоспинальный проводящие пути, кортикоспинальный и спинокортикальный пути здесь переключаются на новые нейроны. На нейронах продолговатого мозга и варолиева моста заканчиваются кортикобульбарные пути.
3. СРЕДНИЙ МОЗГ
Средний мозг — структурно-физиологическое образование ЦНС. Нейроны его объединяются и образуют: четверохолмие, красное ядро, черную субстанцию, ядра глазодвигательного и блокового нервов. Каждому образованию присуща определенная роль.
Четверохолмие. Состоит из передних и задних бугров. Передние бугры получают информацию со зрительных рецепторов и обеспечивают зрительные, ориентировочные и сторожевые рефлексы, которые выражаются в повороте глаз и головы в сторону действия зрительных раздражителей, повышении тонуса мышц сгибателей конечностей, учащении сокращений сердца, повышении давления крови в сосудах, учащении дыхания (обеспечивают подготовку животного к защите, к бегу, к нападению). Задние бугры воспринимают информацию со слуховых рецепторов и обеспечивают слуховые ориентировочные и сторожевые рефлексы, выражающиеся в настораживании ушей и повороте головы в сторону звука, повышении тонуса мышц сгибателей конечностей, учащении сокращений сердца и дыхания.
Красное ядро. Получает информацию с мозжечка, подкорковых ядер, коры больших полушарий. Участвует в формировании программы действия, которую посылает к нейронам вестибулярного ядра продолговатого мозга, обеспечивающего мышечный тонус. Красное ядро, обеспечивая торможение деятельности мотонейронов, играет большую роль в распределении тонуса мышц, координации двигательных реакций.
Черная субстанция. Взаимосвязана с полосатым телом и бледным шаром. Обеспечивает пластический тонус мышц, участвует в регуляции сложных, точных, тонких двигательных реакций — жевания, глотания, а также вегетативных реакций — дыхания, тонуса сосудов, сокращений сердца.
СТАТИЧЕСКИЕ И СТАТОКИНЕТИЧЕСКИЕ РЕФЛЕКСЫ
ПРОДОЛГОВАТОГО И СРЕДНЕГО МОЗГА
С участием продолговатого и среднего мозга осуществляются перераспределение тонуса мышц в зависимости от положения тела в пространстве, тонические и установочные (выпрямительные), а также статокинетические рефлексы.
Статические тонические или познотонические рефлексы. Обеспечивают поддержание естественной позы животного. Они осуществляются через продолговатый мозг с участием спинного:
1. рефлекс с вестибулярного аппарата на мышцы разгибатели конечностей. Обеспечивает их высокий тонус, сохранение положения позы животного теменем и спиной вверх;
2. рефлекс с вестибулярного аппарата на мышцы сгибатели конечностей. Возникает при положении животного теменем и спиной вниз и проявляется в повышении тонуса мышц сгибателей конечностей;
3. рефлекс с рецепторов мышц шеи на мышцы сгибатели задних конечностей и мышцы разгибатели передних конечностей. Проявляется при запрокидывании головы в выпрямлении передних конечностей и сгибании задних;
4. рефлекс с рецепторов мышц шеи на мышцы разгибатели задних конечностей и сгибатели передних конечностей. Проявляется в сгибании передних и разгибании задних конечностей при наклоне головы и шеи;
5. рефлекс с рецепторов мышц шеи (рефлекс вращения) на мышцы разгибатели конечности одной стороны и мышцы сгибатели противоположной стороны. Проявляется при вращении в разгибании конечностей той стороны тела, в которую поворачивается голова, и в сгибании конечностей противоположной стороны.
Выпрямительные (установочные) рефлексы. Обеспечивают возвращение головы и тела из неестественного положения в естественное. Осуществляются через средний мозг:
1. рефлекс с рецепторов вестибулярного аппарата на мышцы головы возникает при положении головы и туловища на боку. Проявляется в перераспределении тонуса мышц головы и возврате головы в естественное положение;
2. рефлекс с тактильных рецепторов кожи при положении животного лежа на боку на мышцы головы. Обеспечивает возврат головы в естественное положение;
3. рефлекс с рецепторов мышц шеи, возникающий при изменении положения шеи (при повороте ее вокруг своей оси), на мышцы туловища. Обеспечивает перевод туловища в положение, соответствующее положению шеи, за счет перераспределения тонуса мышц;
4. рефлекс с рецепторов кожи туловища, возникающий при положении животного на боку, на мышцы туловища. Обеспечивает за счет перераспределения тонуса мышц возврат туловища в естественное положение, соответствующее положению головы и шеи.
Статокинетические рефлексы. Проявляются при движении животного, изменении положения отдельных частей тела, когда происходит перераспределение тонуса мышц глаз, туловища и конечностей, что обеспечивает устойчивое положение глаз, головы и тела в пространстве:
1. рефлекс с рецепторов мышц одной конечности на мышцы других. Возникает при ходьбе животного, когда при сгибании одной конечности повышается тонус мышц разгибателей других трех конечностей;
2. рефлекс «нистагм головы» возникает при вращательных движениях. Проявляется в движении головы в противоположную сторону вращения туловища, а затем в скачкообразном перемещении головы в положение, соответствующее положению туловища;
3. рефлекс «нистагм глаз» возникает при вращательных движениях. Проявляется в движении глаз в противоположную сторону вращения головы и туловища, а затем в скачкообразном перемещении глаз в положение, соответствующее положению туловища;
4. лифтовый рефлекс возникает при линейном ускорении движения вверх и вниз; в первом случае повышается тонус мышц разгибателей, во втором — тонус мышц сгибателей.
4. МОЗЖЕЧОК
Мозжечок — структурно-физиологическое образование ЦНС. Нейроны его объединяются и образуют ядра мозжечка, поверхностный слой, или кору. Анатомически он состоит из двух полушарий и средней части, которая их соединяет. Ядра мозжечка связаны проводящими путями (имеют прямые и обратные связи) с корой больших полушарий, средним и продолговатым (красным ядром и вестибулярными ядрами) и спинным (мотонейронами) мозгом. В связи с этим в мозжечке различают три зоны: корковую, вестибулярную и спинальную. Нейроны мозжечка имеют прямые и обратные связи с ретикулярной формацией.
Мозжечок получает информацию с рецепторов мышц, глаз, вестибулярного аппарата, с коры больших полушарий. Через ретикулярную формацию, красное ядро, ядро Дейтерса он связан с мотонейронами спинного мозга. Участвует в обеспечении тонуса мышц, позы, координации движений, оптимальной возбудимости и лабильности вегетативных и соматических центров, равновесия тела при движении.
Удаление мозжечка вызывает атонию (исчезновение тонуса), астению (быстрое утомление), атаксию (шаткость походки), астазию (дрожащие движения).
5. РЕТИКУЛЯРНАЯ ФОРМАЦИЯ
Ретикулярная, или сетчатая формация, представляет собой самостоятельное структурно-физиологическое образование ЦНС, которое расположено главным образом в продолговатом и среднем мозге. Нейроны ее имеют короткие и ветвистые отростки, которые переплетаясь, образуют подобие сети. Нейроны объединяются в ядра. Отростки нейронов ретикулярной формации идут к различным отделам ЦНС и образуют восходящую и нисходящую системы.
Восходящая система образована нейронами и отростками их, связанными с корой больших полушарий; нисходящая система — с мозжечком, красным ядром, мотонейронами спинного мозга, нейронами симпатического отдела вегетативной нервной системы. Ретикулярная формация через симпатическую нервную систему осуществляет облегчающие и тормозящие деятельность влияния на все нервные проводники, рецепторы и все внутренние органы, мышцы. Ретикулярная формация оказывает свое влияние на все нервные центры.
Ретикулярная формация активируется потоками импульсов, поступающими к ней со всех рецепторов организма по неспецифическим путям, с мозжечка, коры больших полушарий, таламуса, лимбической системы, красного и вестибулярного ядер. Получив информацию, она формирует свою программу и передает ее на восходящую и нисходящую системы.
По восходящим путям программа поступает к нейронам коры больших полушарий, вызывает и поддерживает некоторое постоянное возбуждение их, т. е. поддерживает тонус коры больших полушарий. Поддержание тонуса коры больших полушарий имеет большую физиологическую значимость, так как только в этом случае кора может осуществлять свою специфическую деятельность — воспринимать информацию и отвечать на нее. Ретикулярная формация обеспечивает состояние бодрствования и сна, участвует в расшифровке поступающей информации с рецепторов путем регуляции потока импульсов.
По нисходящим путям программа передается к нервным центрам, нервным проводникам, рецепторам, органам и обеспечивает повышение или понижение их возбудимости и тем самым оптимальную деятельность.
6. ПРОМЕЖУТОЧНЫЙ МОЗГ
Промежуточный мозг представляет собой самостоятельное структурно-физиологическое образование ЦНС, нейроны которого имеют большую физиологическую значимость в нервных центрах. В нем выделяют три основных самостоятельных структуры: таламус, или зрительные бугры, гипоталамус, или подбугровая область, и эпиталамус, или надталамическая область, — свод и эпифиз.
Таламус, или зрительные бугры. Представляет собой скопление ядер, образованных нейронами. Все ядра таламуса по физиологической значимости делят на специфические, ассоциативные, моторные и неспецифические.
Специфические ядра таламуса имеют двухсторонние прямые связи с определенными участками коры больших полушарий. Они получают информацию со всех рецепторов организма (исключение — обонятельные рецепторы), подвергают ее первичному анализу и переключают на пути к коре больших полушарий. Таким образом, благодаря циркуляции информации с рецепторов между специфическими ядрами таламуса и сенсорными нейронами коры больших полушарий происходит анализ (определение биологической значимости поступающей информации), синтез (формирование программы действия) и обеспечивается целостное восприятие поступающей информации.
Нейроны ядер таламуса связаны с нейронами гипоталамуса, участвующими в регуляции деятельности внутренних органов и
мышц.
Ассоциативные ядра таламуса получают информацию от специфических ядер. Они связаны с нейронами коры больших полушарий и участвуют в интеграции деятельности различных образований мозга.
Моторные ядра таламуса получают информацию от мозжечка и базальных ганглиев. Посылают информацию в моторную зону коры больших полушарий и участвуют в регуляции движений.
Неспецифические ядра таламуса не имеют прямых связей с конкретными участками коры больших полушарий. Они образуют широкие взаимные связи со специфическими ядрами таламуса и получают информацию с них. Получив информацию, они рождают собственные импульсы и передают их в ту область коры больших полушарий, в которую в данный момент поступает специфическая информация, активируя нейроны коры и повышая их общий тонус. Повышенный тонус нейронов коры — условие для полноценной деятельности их.
Гипоталамус. Образует вентральную часть промежуточного мозга. Состоит из нейронов, которые объединяются в ядра гипоталамуса. Различают преоптическую, переднюю, среднюю, наружную и заднюю группы ядер. Нейроны ядер преоптической группы гипоталамуса продуцируют либерины и статины, регулируют деятельность передней доли гипофиза.
В ядрах гипоталамуса расположены нервные центры. В передних ядрах — высший отдел парасимпатической иннервации, с которого обеспечиваются общие парасимпатические приспособительные реакции; в задних ядрах — высший отдел симпатической иннервации, обеспечивающий симпатические эффекты. В средних ядрах находятся нервные центры регуляции всех видов обмена веществ и энергии, голода и насыщения, терморегуляции, деятельности желез внутренней секреции, половой системы, лактации, почек.
В целом гипоталамус обеспечивает интеграцию деятельности вегетативной, эндокринной и соматической систем; участвует в регуляции поведенческих реакций (половых и агрессивно-оборонительных).
Эпиталамус. Он является железой внутренней секреции. Его называют счетчиком времени. Это своего рода биологические часы.
7. ЛИМБИЧЕСКАЯ СИСТЕМА
Лимбическая система — самостоятельное структурно-физиологическое образование, которое кольцеобразно охватывает основание переднего мозга на границе со стволовой частью мозга. Лимбическая система включает в себя отдельные скопления нейронов: гиппокамп — основная структура системы, поясная извилина, мамилярные тела и др. Она связана с корой больших полушарий, подкорковыми ядрами, таламусом, гипоталамусом и ретикулярной формацией.
Нейроны лимбической системы принимают большую часть информации с различных рецепторных полей тела и внутренних органов. Совместно с корой больших полушарий, подкорковыми ядрами, таламусом, ретикулярной формацией участвует в анализе и синтезе ее, формировании программы действия, которую передают на исполнительные органы через гипоталамус, обеспечивая постоянство условий внутренней среды организма, вегетативные реакции (ее называют висцеральным мозгом). Лимбическая система участвует в механизмах памяти, контроле активности мозга (бодрствования и сна), в формировании эмоциональной окраски поведения животных.
8. ПОДКОРКОВЫЕ ЯДРА
Подкорковые (базальные) ядра, образованные нейронами, располагаются в белом веществе больших полушарий головного мозга. Они представляют собой самостоятельные структурно-физиологические образования. Наиболее изучены из них: хвостатое ядро, скорлупа и бледный шар, называемые стриопаллидум (стриатум + паллидум). Он имеет обширные связи с другими отделами центральной нервной системы.
Паллидум, или бледный шар. Важный отдел нервного центра, обеспечивающий согласованную деятельность всех мышц туловища. Он образован большими нейронами. Получая афферентную информацию с полосатого тела и рецепторов скелетных мышц, он совместно со спинным, продолговатым, средним мозгом, мозжечком, ретикулярной формацией, таламусом и корой больших полушарий формирует программу действия, обеспечивающую согласованную деятельность всех мышц туловища при сложных двигательных реакциях (при беге, жевании, нападении и др.).
Стриатум, или полосатое тело. Включает в себя хвостатое ядро и скорлупу, образовано мелкими нейронами. Стриатум получает афферентную информацию с сенсорной зоны коры больших полушарий и черной субстанции среднего мозга. Аксоны нейронов полосатого тела направляются к бледному шару и черной субстанции. Аксоны нейронов бледного шара направляются к ядрам промежуточного и среднего мозга. От ядер таламуса эфферентный путь идет к двигательным нейронам коры. Благодаря циркуляции информации по этим связям формируется программа действия, обеспечивающая согласованную деятельность мышц туловища и внутренних органов, целенаправленные движения.
9. КОРА БОЛЬШИХ ПОЛУШАРИЙ ГОЛОВНОГО МОЗГА
Кора больших полушарий головного мозга — это наиболее развитый отдел головного мозга, который покрывает полушария снаружи. Она представляет собой тонкий слой серого вещества. Полушария состоят из белого вещества. Толщина коры 1,5...3 мм; 6 слоев — I...VI (рис. 3).
Нейроны коры отличаются по форме (в основном пирамидные, веретенообразные и звездчатые) и имеют множество типов соединений. Они выполняют разные роли.

Рис.3. Расположение нейронов и нервных волокон в коре больших полушарий:
I— молекулярный слой, образован главным образом сплетением нервных волокон, нервных клеток незначительное число;
II— наружный зернистый слой, образован плотно расположенными мелкими овальной, треугольной или многоугольной формы нейронами;
III — слой пирамидных нейронов разных размеров; IV — внутренний зернистый слой, образован скоплением мелких овальной, треугольной или многоугольной формы нейронов; V —слой гигантских пирамидных клеток; VI — мультиформенный слой, содержит нейроны веретенообразной и треугольной формы
По функциональному значению все нейроны коры больших полушарий делят на три группы:
1) чувствительные (сенсорные и ассоциативные) — обеспечивают восприятие импульсов непосредственно с рецепторов (сенсорные) и от ядер таламуса, а через него от различных рецепторных полей (ассоциативные);
2) моторные — посылают импульсы от коры к нижележащим структурам ЦНС и рабочим органам, являющиеся представителями нервных центров безусловных рефлексов в коре больших полушарий;
3) контактные — осуществляют связь между нейронами коры больших полушарий.
Чувствительные нейроны. Расположены в III и IV слоях коры и образуют воспринимающие зоны: сенсорные и окружающие их ассоциативные.
Сенсорные нейроны. Образуют сенсорные зоны. Каждая зона называется соответственно рецепции, в которой она участвует (рис. 4). Размеры каждой сенсорной зоны зависят от физиологической значимости для организма животного рецепции. Чем выше значимость, тем она больше.
Выделяют следующие сенсорные проекционные зоны:
1) двигательная — расположена между лобной и теменной долями. Раздражение этой зоны вызывает сокращение мышц. Рядом с ней находится вторичная двигательная зона. Эти зоны имеют и сенсорные входы. Поэтому их называют первичная и вторичная мотосенсорные зоны',

Рис.4. Схема расположения проекционных сенсорных зон в коре больших полушарий собаки:
темные кружки — сенсорные клетки зрительной зоны; белые кружки — сенсорные клетки чувствительно-двигательной зоны; треугольники — сенсорные клетки слуховой зоны; квадраты — сенсорные клетки вкусовой зоны; ромбы — сенсорные клетки двигательной зоны
2) соматосенсорная первичная и вторичная (или чувствительно- двигательная) — расположена в лобной и теменной долях, вдоль центральной борозды; воспринимает импульсы с рецепторов кожи и двигательного аппарата через таламус;
3) слуховая — расположена в височной доле; воспринимает импульсы от слуховых рецепторов;
4) зрительная — лежит в затылочной области; воспринимает импульсы от рецепторов сетчатки глаз;
5) обонятельная и 6) вкусовая — лежат на внутренней поверхности коры; связаны, соответственно, с обонятельными рецепторами носовых раковин и вкусовыми рецепторами языка и ротовой полости. Имеют двусторонние связи с лимбической системой.
Ассоциативные зоны располагаются рядом с проекционными зонами. Их нейроны участвуют в анализе информации, в осуществлении связи между сенсорными и двигательными нейронами. Без ассоциативных нейронов невозможен четкий анализ и синтез программы (например, животное не откликается на свою кличку после удаления ассоциативной зоны в слуховой сенсорной зоне).
Моторные нейроны. Располагаются в V слое коры больших полушарий, образуют в ней корковые отделы нервных центров безусловных рефлексов. Моторные нейроны объединяются группами и образуют моторные зоны. Каждая моторная зона обеспечивает связь коры с органами организма. Моторные зоны способны переводить органы из состояния покоя в деятельное.
Контактные нейроны. Осуществляют связь между разными нейронами в коре больших полушарий.
В коре имеется большое количество глиальных клеток (в 10 раз больше, чем нейронов), выполняющих опорную, обменную, секреторную (выделяют вещества, регулирующие возбудимость нейронов) роли, а также участвующих в хранении следов осуществленных реакций (память).
Любая ответная реакция связана с работой ряда различных зон, составляющих так называемую распределительную систему.
Нейроны коры больших полушарий находятся в состоянии тонуса (постоянного возбуждения), который не исчезает и во время сна. Показателем постоянного тонуса нейронов коры являются биотоки, которые могут быть зарегистрированы в виде электроэнцефалограммы.
Наряду с проекционными зонами, связанными с выполнением сенсорной и моторной ролей, в пределах теменной, височной и лобной долей расположены поля, составляющие ассоциативную кору, для нейронов которой свойственно отвечать на раздражения различной модальности и таким образом участвовать в интеграции сенсорной информации и в обеспечении связи между сенсорными и двигательными зонами коры. Они участвуют в оценке биологически значимой информации и в восприятии пространственных отношений окружающего мира, контролируют оценку мотивационного поведения и программирование сложных поведенческих актов.
Соматосенсорная и другие сенсорные зоны, мотосенсорные и сенсомоторные зоны коры организованы в элементарные функциональные единицы — колонки, в которых осуществляется переработка информации от рецепторов одной модальности. Каждая колонка состоит из нескольких микромодулей, объединяющих 5 (6) гнездообразно расположенных пирамидных, звездчатых, верете-новидных нейронов. Согласованная деятельность их и обеспечивает формирование полноценных программ действия, приспособительных реакций.
В целом кора больших полушарий совместно с подкорковыми образованиями осуществляет свою деятельность по принципу рефлекса. В отличие от других структурно-физиологических образований ЦНС осуществляет свою работу по принципу условного рефлекса, поэтому ее деятельность называется условно-рефлекторной, или высшей нервной деятельностью. Характерная особенность условных рефлексов заключается в том, что они образуются в течение индивидуальной жизни организма. Высшая нервная деятельность связана с явлениями психической жизни животных и человека, обеспечивает целесообразность поведения в меняющихся условиях: запоминание полезных признаков, способность приобретать жизненный опыт, обучение.
10.ПЕРИФЕРИЧЕСКИЙ СОМАТИЧЕСКИЙ ОТДЕЛ НЕРВНОЙ СИСТЕМЫ
Периферический соматический отдел нервной системы обеспечивает осуществление ЦНС двигательных реакций. Соматические нервы образованы аксонами мотонейронов и аксоноподобными дендритами рецепторных нейронов, которые идут из ЦНС, спинномозговых и черепно-мозговых ганглиев к периферическим исполнительным органам не прерываясь; скорость проведения возбуждения большая и составляет 30... 120 м/с.
Соматический отдел нервной системы представлен черепно-мозговыми и спинномозговыми нервами.
Черепно-мозговые нервы. От головного мозга отходят 12 пар нервов. Они являются смешанными; содержат аксоны нейронов головного мозга и рецепторные нейроны, или только первые, образующие эфферентные нервы, или только вторые, образующие афферентные нервы за счет своих отростков. Они включаются в рефлекторные дуги соматических и вегетативных рефлексов.
По черепно-мозговым нервам происходит передача программ действия к мышцам головы, внутренним органам, поступление информации в ЦНС с рецепторов кожи головы, зрительных, слуховых, обонятельных и вкусовых рецепторов, интерорецепторов.
Спинномозговые нервы. Отходят симметричными парами по обе стороны спинного мозга. Через дорсальные корешки в спинной мозг входят аксоны рецепторных нейронов, через вентральные - выходят из спинного мозга аксоны двигательных нейронов; эти отростки, объединяясь, образуют спинномозговые нервы.
По спинномозговым нервам передается программа действия на мышцы туловища и конечностей, поступает информация в ЦНС с экстеро- и интерорецепторов.
11.ВЕГЕТАТИВНЫЙ ОТДЕЛ НЕРВНОЙ СИСТЕМЫ
Вегетативный отдел нервной системы представлен парасимпатической, симпатической и метасимпатической (энтеральной) иннервацией, каждая из которых имеет ряд особенностей (рис. 5).
Вегетативная нервная система состоит из вегетативных нейронов, расположенных в среднем, продолговатом и спинном мозге, а также в ганглиях на периферии. Для нее характерен двухнейронный принцип образования.
Центральную часть вегетативной нервной системы составляют первые нейроны, расположенные в среднем, продолговатом и спинном мозге.
Периферическое звено парасимпатической и симпатической иннервации представляет собой цепь из двух последовательно соединенных нейронов. Аксоны первых нейронов (нервные волокна) выходят из ЦНС и заканчиваются обязательно на вторых нейронах, объединенных в ганглии (узлы). Аксоны вторых нейронов идут к иннервируемому органу. Скорость проведения возбуждения по вегетативным нервным волокнам составляет 2...14 м/с.
К периферической части относят и висцеральные афференты, т. е. чувствительные нервные волокна, проходящие в составе блуждающих, языкоглоточных и чревных нервов. Тела нейронов, к которым идут эти волокна, располагаются в соответствующих ганглиях названных нервов и спинномозговых узлах.
Вегетативный отдел нервной системы обеспечивает регуляцию структурной организации и деятельности внутренних органов, сосудов, потовых желез, а также трофику (питание) всех структур, включая скелетные мышцы, рецепторов и самой нервной системы.
Высшие нервные центры вегетативного отдела нервной системы находятся в гипоталамусе: в передних ядрах — центры парасимпатической иннервации, в задних ядрах — центры симпатической иннервации.

Рис.5. Принципиальная схема дуги вегетативного рефлекса:1 — спинной мозг; 2 — чувствительный нейрон; 3 — спинномозговой ганглий; 4 — дорсальный корешок, 5 — преганглионарный нейрон дуги вегетативного рефлекса; 6 — вентральный корешок; 7 — преганглио-нарное нервное волокно; 8 — симпатический паравертебральный ганглий (узел симпатического ствола); 9 — белая соединительная
Парасимпатическая иннервация. Образована парасимпатическими нейронами среднего, продолговатого мозга и крестцового от дела спинного мозга, а также парасимпатическими нейронами ганглий, расположенных чаще в органах. Аксоны нейронов среднего мозга направляются к исполнительным органам в составе глазодвигательного нерва, продолговатого мозга — в составе лицевого, языкоглоточного и блуждающего нервов; крестцового отдела спинного мозга — в составе тазовых нервов. Они называются преганглионарными парасимпатическими волокнами. Ганглии лежат около органа или прямо в иннервируемом органе. Передача возбуждения с аксонов первых нейронов (преганглионарных волокон) на нейроны ганглий и с аксонов этих нейронов (постганглионарных волокон) на структуры органа осуществляется через синапсы с помощью медиатора ацетилхолина. Преганглионарное волокно длинное, так как идет от ЦНС до органа; постганглионарное — короткое.
Симпатическая иннервация. Образована симпатическими нейронами, расположенными в боковых рогах грудного и поясничного отделов спинного мозга (от 1-го грудного до 2...4-го поясничного), а также симпатическими нейронами ганглий, расположенными по обе стороны около грудных и поясничных позвонков (вертебральные ганглии) или вдали от позвонков (превертебральные ганглии: краниальный и каудальный шейные, звездчатый, чревный и краниальный брыжеечный, объединяются как солнечное сплетение, каудальный брыжеечный). Аксоны симпатических нейронов спинного мозга (преганглионарные волокна) направляются на периферию через вентральные корешки спинного мозга и оканчиваются на нейронах вертебральных или превертебральных узлов. Образуют пограничные симпатические стволы, идущие по вентральной поверхности позвонков. Аксоны нейронов этих узлов (постганглионарные волокна) идут к периферическим исполнительным органам и оканчиваются на них. Образуют крупные периферические симпатические нервы: грудные, внутренностные, малые и большие чревные, иннервирующие соответственно органы грудной и брюшной полостей, сосуды. Симпатические нервные волокна включаются и в соматические нервы, направляются к сосудам мышц, рецепторам кожи и мышц. Передача возбуждения с преганглионарного волокна на постганглионарное осуществляется через синапсы с помощью медиатора ацетилхолина и холинорецепторов. Преганглионарные волокна короткие, постганглионарные длинные. Передача импульсов с постганглионарного волокна на орган осуществляется через синапсы с помощью медиатора норадреналина и адренорецепторов.
Метасимпатическая (энтеральная) иннервация. Большинство полых висцеральных органов (сердце, бронхи, мочевой пузырь, пищеварительный тракт, матка, желчный пузырь, желчные пути и др.) наряду с симпатической и парасимпатической иннервацией имеют собственный местный механизм регуляции в виде метасимпатической нервной системы. Метасимпатическая нервная система представлена интрамуральными ганглиями, залегающими в толще стенок полых органов, которые изолированы от окружающих тканей специальными барьерами.
Нейроны интрамуральных ганглий по физиологической роли делятся на чувствительные нейроны, интернейроны, эффекторные нейроны. Нейроны объединяются в рефлекторные дуги. Тела нейронов имеют множество синапсов; отростки нервных клеток содержат большое количество пузырьков с медиаторами. Передача возбуждения в нейронах, составляющих ганглии метасимпатической системы, осуществляется посредством ацетилхолина и норадреналина. В синапсах постганглионарных волокон выделяются разнообразные вещества — ацетилхолин, норадреналин, АТФ, аденозин, серотонин, дофамин, адреналин, гистамин и др. Однако главная роль в передаче возбуждения в метасимпатической системе принадлежит АТФ и аденозину. Воспринимающие АТФ и аденозин рецепторы называются пуринергтескими.
Метасимпатическая нервная система иннервирует только внутренние органы.
12.ПРИНЦИП ДЕЯТЕЛЬНОСТИ ВЕГЕТАТИВНОГО ОТДЕЛА НЕРВНОЙ СИСТЕМЫ
Вегетативный отдел нервной системы осуществляет свою деятельность по принципу безусловных и условных вегетативных рефлексов.
Афферентный путь рефлекторной дуги вегетативного рефлекса представлен висцеральными (висцеральные афференты) и соматическими нервными волокнами. Вегетативные нервные центры заложены в спинном и продолговатом мозге, высшие центры — в гипоталамусе. Эфферентный путь — периферическое звено вегетативной нервной системы (рис. 5).
Число вегетативных рефлексов очень велико и они разнообразны: висцеро-висцеральные, висцеро-кутанные, кутанно-висцеральные рефлексы, висцеро-соматические.
Висцеро-висцеральные — это рефлексы с рецепторов внутренних органов на эти же или другие внутренние органы; висцеро-кутанные — рефлексы с рецепторов внутренних органов на сосуды и другие структуры кожи (рис. 6); кутанно-висцеральные — рефлексы с рецепторов кожи на сосуды и другие структуры внутренних органов.
Через вегетативные нервные волокна осуществляются сосудистые, трофические и функциональные влияния на органы. Сосудистые влияния определяют просвет сосудов, давление крови, кровоток; трофические — проявляются в изменении проницаемости мембран клеток и активности ферментов, обмена веществ в тканях и органах; функциональные — пусковые, коррегирующие, стимулирующие и тормозящие деятельность органов.
Симпатическая иннервация универсальна, так как иннервирует ткани всех органов и кровеносные сосуды скелетных мышц. Парасимпатическая система иннервирует мышцы глаз, слюнные железы, мышцы языка, трахеи и бронхов, легкие, все органы пи-
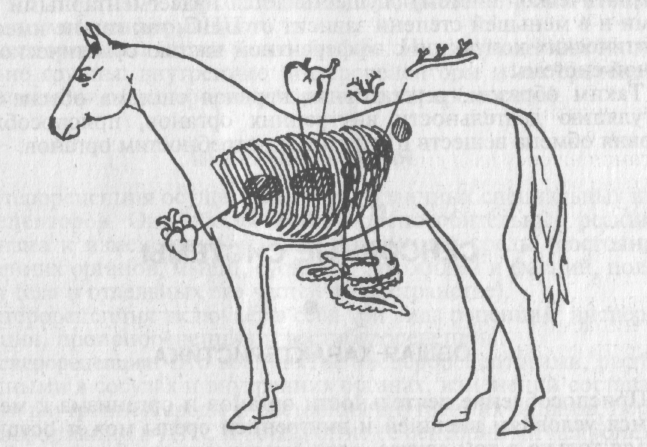
Рис. 6. Висцеро-кутанные рефлексы с рецепторов внутренних органов на сосуды и другие структуры кожи: проявляются в повышении тактильной и болевой чувствительности в зонах Захарьина-Хеда
щеварения, сердце, почки, мочевой пузырь и мочеточники и другие внутренние органы, некоторые кровеносные сосуды (языка, подчелюстной железы, пещеристых тел полового члена). Метасимпатическая нервная система иннервирует только внутренние органы.
Симпатические нервные волокна стимулируют работу сердца (учащают и усиливают сокращения), секреторную деятельность потовых желез, обмен веществ в мышцах, суживают кровеносные сосуды, совместно с парасимпатической иннервацией обеспечивают деятельность пищеварительного аппарата (ослабляют сокоотделение и тормозят моторику), расширяют зрачки, расслабляют стенку мочевого пузыря и др. Деятельность ее активируется при действии на организм неблагоприятных факторов.
Парасимпатические нервные волокна совместно с симпатическими обеспечивают оптимальную деятельность органов, которые они иннервируют, при стрессовых воздействиях оказывают чаще влияния, противоположные действию симпатической нервной системы.
Метасимпатическая нервная система программирует и координирует двигательную, секреторную и всасывательную активность органов, активность местных эндокринных элементов и локальный кровоток. Она определяет способности органов ритмически сокращаться с определенной частотой и амплитудой без воздействия извне под влиянием метаболических изменений в самом органе. Координация ее деятельности (как симпатической и парасимпатической систем) осуществляется надсегментарными центрами и в меньшей степени зависит от ЦНС, так как не имеет синаптических контактов с эфферентной частью соматической нервной системы.
Таким образом, вегетативная нервная система обеспечивает регуляцию деятельности внутренних органов, приспособление уровня обмена веществ и энергии к потребностям органов.