ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ
ОБЩАЯ ФИЗИОЛОГИЯ ВОЗБУДИМЫХ ТКАНЕЙ. •
ПЛАН ЛЕКЦИИ
1. Законы возбуждения.
2. Биоэлектрические явления.
3. Изменение возбудимости ткани при возбуждении.
4. Физиология мышц.
5. Физиология нервов.
6. Нервно-мышечная передача возбуждения. Синапс.
К возбудимым тканям относятся ткани – нервная, мышечная и железистая. Эти ткани могут находиться в трёх основных состояниях – физиологическом покое, возбуждении и торможении, которые представляют разные стороны их жизнедеятельности. Переход из одного состояния в другое происходит в ходе постепенных количественных изменений.
Физиологическим покоем называется такое состояние, когда ткань или орган не проявляют признаков характерной для них жизнедеятельности. Если мышца не сокращается, значит она находится в состоянии покоя. Состояние покоя условно, т.к. даже в это время в мышце происходит обмен веществ. Деятельное состояние в мышцах и нервах может протекать в виде возбуждения и торможения. Возбуждением называется деятельное состояние живой ткани, в которое она приходит под влиянием раздражителя. Под раздражителем понимают любое изменение условий внешней и внутренней среды, если оно возникает внезапно, имеет достаточную силу, удерживается определенное время, вызывает обратимые изменения структуры и деятельности живых тканей и клеток. Процесс воздействия раздражителя на живые структуры называется раздражением.
По происхождению раздражители бывают: физические (термические, механические, электрические, световые, звуковые), физико-химические (изменение осмотического давления, реакция среды, электролитный состав), химические (вещества пищи, гормоны, продукты обмена, яды, лекарственные вещества, кислоты, щелочи) и биологические. Особым раздражителем является нервный импульс.
По физиологическому значению раздражители подразделяют на адекватные и неадекватные. Адекватные - это раздражители, к которым организм и его структуры в естественных условиях в процессе эволюции приспособились к восприятию этого раздражителя (например, свет адекватен для структур глаза). Неадекватные - это раздражители, которые в естественных условиях не действуют на организм, и структуры организма не приспособлены к их восприятию. Поэтому такие раздражители чаще всего вызывают нарушение функции организма.
По силе – раздражители бывают допороговые, пороговые и сверхпороговые. Минимальная сила раздражения, способная вызвать ответную реакцию, называется порогом раздражения, а раздражители, не способные вызвать ответную реакцию, называются подпороговыми. Сверхпороговые – это раздражители, сила которых выше пороговой. В результате воздействия на ткань очень мощных (запредельных) раздражителей ответная реакция может не наступить.
Возбудимые ткани обладают рядом специфических свойств: возбудимостью и проводимостью.
Возбудимость - способность возбудимой ткани отвечать изменением структуры и деятельности на действие раздражителя, т.е. отвечать особой биологической реакцией, называемой возбуждением.
Возбуждение - ответная реакция возбудимой ткани на действие возбудителя (раздражителя), проявляющаяся в совокупности физических, физико-химических, химических, метаболических процессов и изменений деятельности. Возбуждение - волнообразный процесс, который проявляется в разных возбудимых тканях специфическим образом: в мышечной - сокращением, в железистой - образованием и выделением секрета, в нервной -возникновением и проведением нервного импульса.
Развитие возбуждения сопровождается кратковременным (в нервном волокне 0,5мс) исчезновением возбудимости (абсолютная рефрактерность). Затем она быстро (4 мс) восстанавливается (относительная рефрактерность).
Обязательным и общим признаком возбуждения возбудимых тканей является возникновение биологического тока действия, т.е. биоэлектрических явлений.
Проводимость - это свойство возбудимой ткани активно проводить волну возбуждения. Например, двигательный нерв кошки проводит возбуждение со скоростью 1200 см/с.
ЗАКОНЫ ВОЗБУЖДЕНИЯ
Ткани и органы организма реагируют на действие раздражителя по определенным законам раздражения.
Первый закон - закон силы. Ткань отвечает на действие раздражителя только в том случае, если раздражитель имеет определенную силу. Минимальная сила раздражителя, способная вызвать возбуждение ткани, называется пороговой силой раздражителя, сила ниже пороговой называется подпороговой, а выше - сверхпороговой. Если в качестве раздражителя используется электрический ток, то минимальная сила электрического тока, способная вызвать возбуждение ткани, называется реобазой.

Исходя из этого закона, для перехода возбудимой ткани из состояния покоя в состояние возбуждения необходимо, чтобы сила действующего раздражителя достигла пороговой (критической) величины. Чем возбудимее ткань, тем меньше для нее пороговая сила возбудителя и, следовательно, более слабый раздражитель может вызвать возбуждение. Например, возбудимость мышцы ниже, чем нерва.
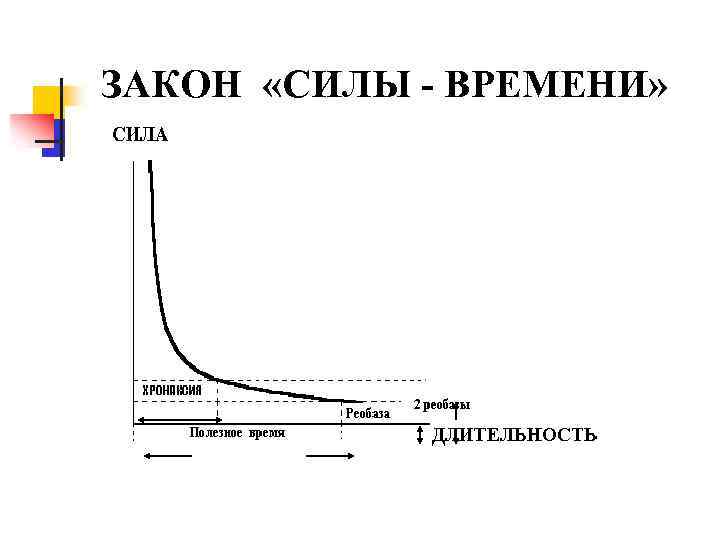
Второй закон - закон времени. Ткань отвечает на действие раздражителя пороговой силы и выше только в том случае, если раздражитель действует определенное время, которое для разных тканей неодинаково. Наименьшее время действия раздражителя пороговой силы, необходимое для того, чтобы вызвать возбуждение, называется полезным временем. Это очень малая величина, непостоянная и трудно определяемая. Она изменяется в связи с естественными изменениями возбудимости ткани под действием целого ряда факторов. В связи с этим для оценки возбудимости ткани было предложено определять хронаксию. Хронаксия - это наименьшее время действия удвоенной реобазы; измеряется в миллисекундах (мс).
Полезное время и хронаксия являются мерой возбудимости ткани; чем они меньше, тем выше возбудимость ткани, и наоборот. Измерение хронаксии мышц, нервов, железистой ткани широко используют для оценки их функционального состояния.
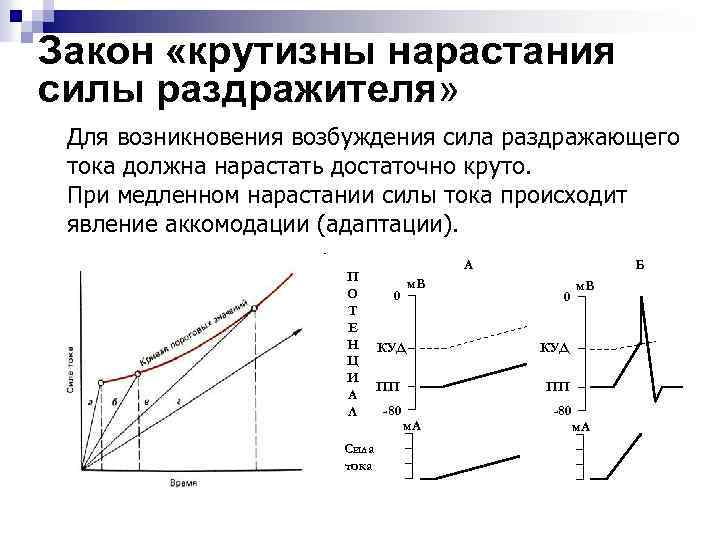
Третий закон - закон крутизны нарастания силы раздражителя. Условием раздражения является нарастание силы с достаточной быстротой, которая характеризуется его крутизной; чем быстрее скорость нарастания силы раздражителя, тем ниже величина пороговой силы, и наоборот. При малой скорости нарастания силы раздражителя, раздражитель может не вызвать ответной реакции ткани. Это связано со свойством ткани приспосабливаться к раздражителю. Изменение состояния тканей, препятствующее развитию возбуждения при длительном действии раздражителя одной силы, называется аккомодацией или приспособлением. Показателем аккомодации является минимальная скорость нарастания силы раздражителя, при которой раздражающий стимул еще способен вызвать ответную реакцию ткани.
Четвертый закон - полярный закон действия раздражителя, или закон действия постоянного тока. При действии постоянного тока на ткань возбуждение возникает только на катоде или аноде, т. е. в момент замыкания цепи постоянного тока возбуждение возникает всегда только под катодом, а в момент размыкания - только под анодом.

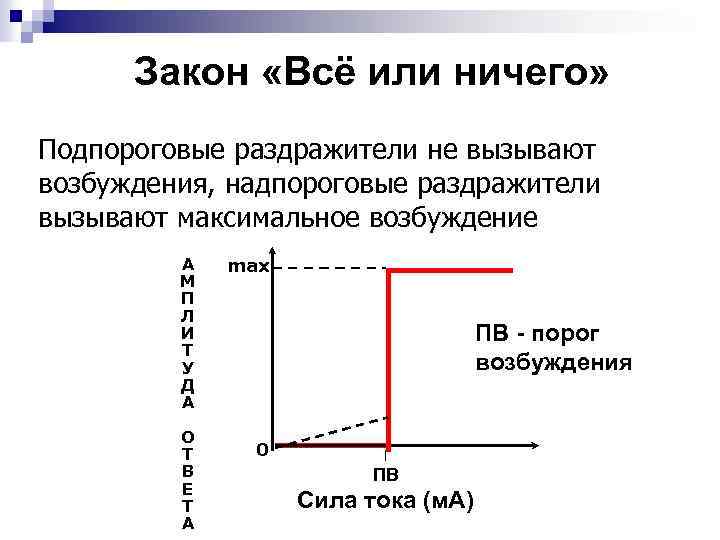
Пятый закон - все или ничего. Структурно-функциональные единицы тканей (клетки, нервные волокна и др.) отвечают на действие возбудителей по закону «все или ничего». На раздражитель подпороговой силы возбудимые ткани не отвечают возбуждением, а на раздражитель пороговой силы отвечают максимальной силы возбуждением. Это универсальный закон, но он не всегда четко проявляется в обычных условиях, потому что ткани состоят из отдельных структурно-функциональных элементов, обладающих разной возбудимостью.
2. БИОЭЛЕКТРИЧЕСКИЕ ЯВЛЕНИЯ
Живая клетка в результате обмена веществ и осуществления своей специфической деятельности непрерывно генерирует электрические потенциалы — биологический ток. По условиям возникновения в живых тканях различают потенциал покоя и потенциал возбуждения, или биологический ток покоя и биологический ток действия.
Между протоплазмой клетки и окружающей клетку средой в живых возбудимых клетках тканей существует ионное неравновесие. В состоянии физиологического покоя внутри клеток больше ионов калия (в 30 - 50 раз), чем снаружи, а снаружи больше ионов натрия (в 10 - 20 раз), чем внутри. Такое ионное неравновесие обеспечивает положительный заряд наружной поверхности и отрицательный заряд внутренней поверхности мембраны клетки, так как большая концентрация ионов натрия (Na+) обеспечивает положительный заряд, а большая концентрация ионов калия (К+) - отрицательный заряд. Если электроды соединить с гальванометром и наложить один электрод на поверхность клетки, а другой ввести внутрь ее, то обнаружится разность потенциалов (биологический ток), равная 15...90 милливольт (мВ). Ток, регистрируемый в возбудимых тканях в состоянии покоя, называется биологическим током покоя, или потенциалом покоя.
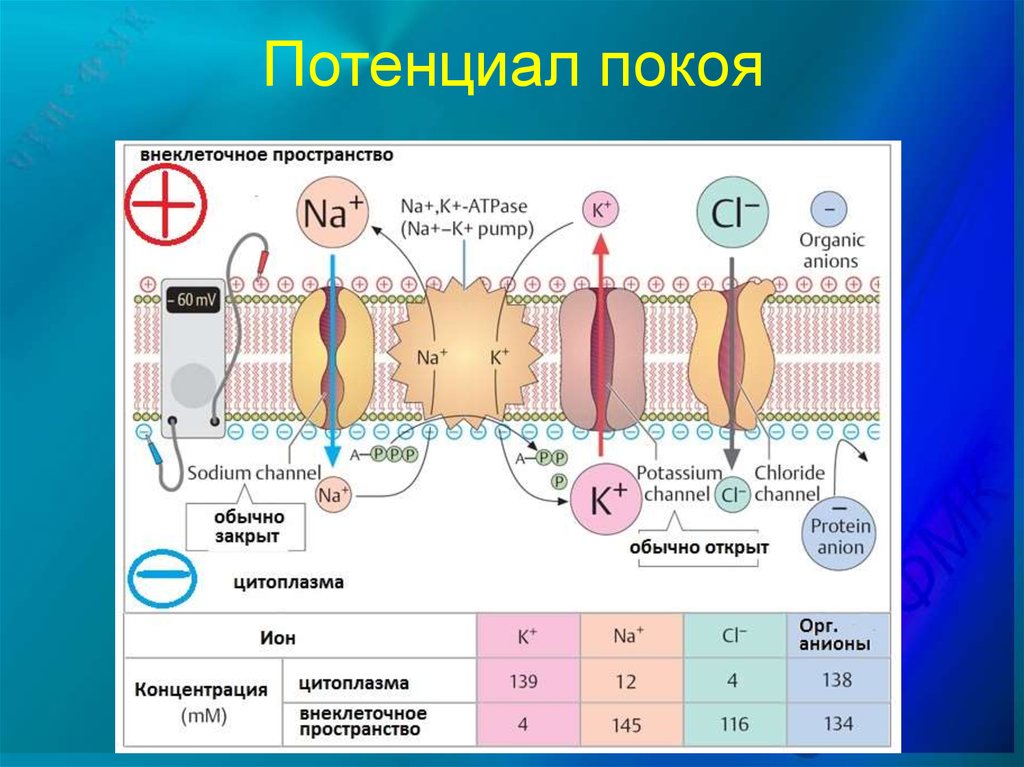
При действии раздражителя на ткань происходит колебание потенциала покоя, а возникающий в этих условиях ток называется током действия, или потенциалом действия. Причиной его возникновения является изменение ионной проницаемости мембраны в участке, на который действует раздражитель: увеличивается поступление ионов натрия во внутрь, а ионов калия - наружу клетки. Это ведет к тому, что поверхность мембраны клетки на месте раздражения становится электроотрицательной, создается разность потенциалов между соседними участками поверхности мембраны клетки, возникает биологический ток, который бежит по мембране клетки. Это и есть биологический ток действия, или потенциал действия. Изменение электрического состояния клеточной мембраны является обязательным признаком возбуждения.

Под влиянием раздражения заряд мембраны начинает быстро уменьшаться от нуля и затем вновь появляться, но только с обратным знаком. Происходит перезарядка, или реверсия, заряда мембраны: внутренняя её сторона заряжается положительно, а наружная – отрицательно. Затем вновь восстанавливаются первоначальные заряды мембраны. Пикообразное колебание потенциала, возникающее в результате кратковременной перезарядки мембраны и последующего восстановления её исходного заряда и называется потенциалом действия. Такое колебание потенциала продолжается 1-5 мс. Величина ПД нерва и скелетной мышцы составляет 115-120 мВ. ПД возникает только под влиянием порогового раздражителя, если сила раздражителя меньше пороговой - возникает местный потенциал, который не приводит к появлению ПД.
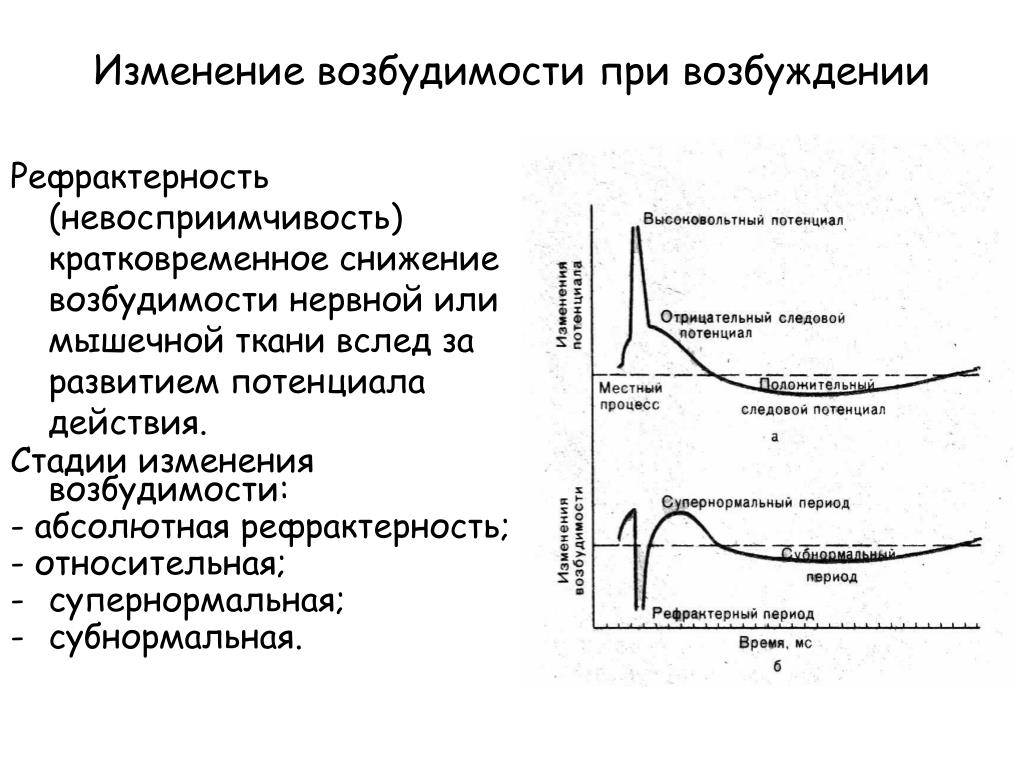
В ПД различают пик длительностью 0,5-0,8 мс и следовые потенциалы. Пик ПД связан с процессом возбуждения, он имеет восходящую и нисходящую фазы. Изменение исходной поляризации мембраны во время восходящей фазы называют деполяризацией, смену знака потенциала называют реверсией (овермут). Вершина пика ПД называется спайк. Во время нисходящей фазы заряд мембраны возвращается к уровню покоя (реполяризация). При реполяризации отмечают следовые потенциалы: следовую деполяризацию и следовую гиперполяризацию.
3. Изменения возбудимости ткани при возбуждении.
Возникновение и развитие импульсов возбуждения сопровождаются последовательными фазовыми изменениями возбудимости ткани. В период развития деполяризации (местный деполяризационный пороговый потенциал) и появления местного нераспространяющегося возбуждения происходит небольшое, очень короткое по времени повышение возбудимости. При переходе местного возбуждения в распространяющееся возникает пик потенциала действия и возбудимость ткани резко падает. Состояние ткани, когда она после раздражения временно не реагирует на повторное раздражение любой силы, называется абсолютной рефракторностью (от лат. refracta — невосприимчивость). В эту фазу повторное раздражение не способно вызвать новый потенциал действия. Абсолютная рефракторность совпадает с восходящей частью пика потенциала действия и длится в мякотных нервных волокнах теплокровных животных примерно 0,5-1 мс, в скелетных мышцах - 2,5-3, а в сердечной мышце -300-400 мс.
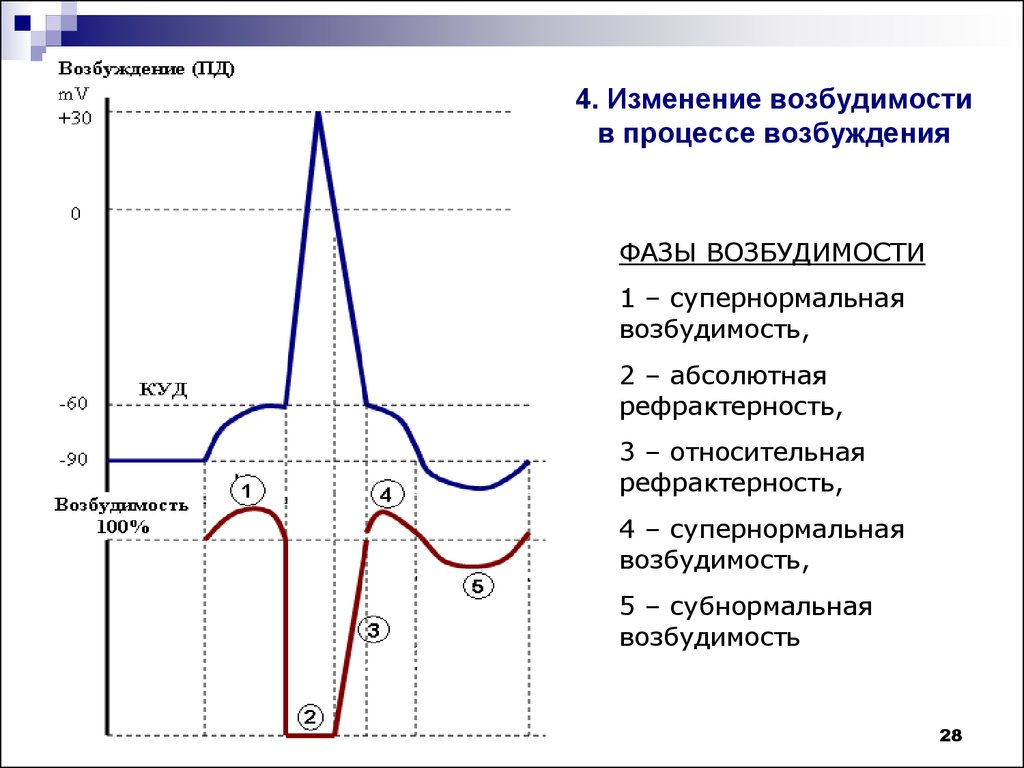
После абсолютной рефракторности возбудимость ткани постепенно восстанавливается до исходного уровня. Период пониженной возбудимости получил название относительной рефракторности. Эта фаза совпадает с реполяризацией, то есть с нисходящей частью пика потенциала и переходом его в следовую деполяризацию. Длительность относительной рефракторности в нервных волокнах составляет от 1 до 10, а в мышцах - до 30 мс.
Затем наступает период повышенной возбудимости, названный фазой экзальтации (от лат. exsaltatio -очень возбужденный). Длительность этой фазы в нерве - 20, в мышце - 50 мс. Она совпадает по времени с периодом окончания следовой деполяризации.
За фазой экзальтации идет длительная фаза субнормальности, когда возбудимость ткани незначительно снижена по сравнению с величиной возбудимости в состоянии физиологического покоя. Эта фаза совпадает со следовой гиперполяризацией. Таким образом, изменениям потенциала действия соответствуют фазы изменения возбудимости.
Абсолютная рефракторность обусловлена тем, что вслед за повышенной проницаемостью мембраны для ионов натрия, приводящей к возникновению потенциала действия, начинается процесс инактивации механизма натриевой проницаемости и в то же время возрастает проницаемость мембраны для ионов калия. В результате этих процессов мембрана временно утрачивает способность возбуждаться и генерировать потенциал действия на новые раздражения. В фазу относительной рефракторности постепенно устраняется инактивация натриевой проницаемости и уменьшается повышенная проницаемость для калия. Поэтому постепенно восстанавливается способность мышечной и нервной тканей возбуждаться и генерировать потенциал действия. Но для возникновения возбуждения требуется раздражитель значительной силы, так как возбудимость клетки еще остается пониженной по сравнению с исходным уровнем. В период относительной рефракторности в возбужденной ткани развиваются процессы, направленные на восстановление исходных свойств, характерных для состояния покоя. При условиях, неблагоприятных для обмена веществ, восстановительные процессы замедляются и удлиняется фаза относительной рефракторности.
В фазу экзальтации восстановительные процессы в клетке заканчиваются. Во время следовой деполяризации потенциал приближается к уровню критической деполяризации и возбудимость клетки повышается. Когда новый раздражитель застает клетку в этом состоянии, то возрастает его эффект, хотя сила раздражителя не изменилась. Поэтому даже допороговый раздражитель будет действовать как сверхпороговый. Во время фазы экзальтации ткань подготовлена для повторного возбуждения. Наличие данной фазы указывает, что раздражитель не только вызывает непосредственную реакцию ткани, но и служит также средством создания наиболее благоприятных условий для новых повторных импульсов возбуждения. Фаза экзальтации играет важную физиологическую роль в осуществлении ритмической деятельности нервной и мышечной тканей. Когда раздражение наносится в ритме, совпадающем по времени с фазой экзальтации, тогда обеспечивается наиболее эффективная деятельность ткани.
Фаза субнормальности возникает в тот период, когда имеется следовая гиперполяризация. При этом потенциал покоя увеличен и, чтобы вызвать деполяризацию до необходимого критического уровня, требуется нанести раздражитель большей силы.
Восстановление ионного неравновесия в клетках тканей обеспечивает специальная система, которая называется калий-натриевый насос. Он представлен специальными переносчиками ионов калия и натрия, которые транспортируют ионы калия внутрь клеток, а ионы натрия из клетки во внешнюю среду и восстанавливают ионное неравновесие в клетке. Переносчиками служат белки-ферменты, локализованные в мембране клеток.
Функциональная подвижность - лабильность. Физиологическое состояние мышечной и нервной тканей, кроме возбудимости, характеризуется также свойством функциональной подвижности, или лабильности. Свойство лабильности открыл Н. Е. Введенский (1892), изучая действие ритмических раздражений различной частоты на нервно-мышечный препарат.
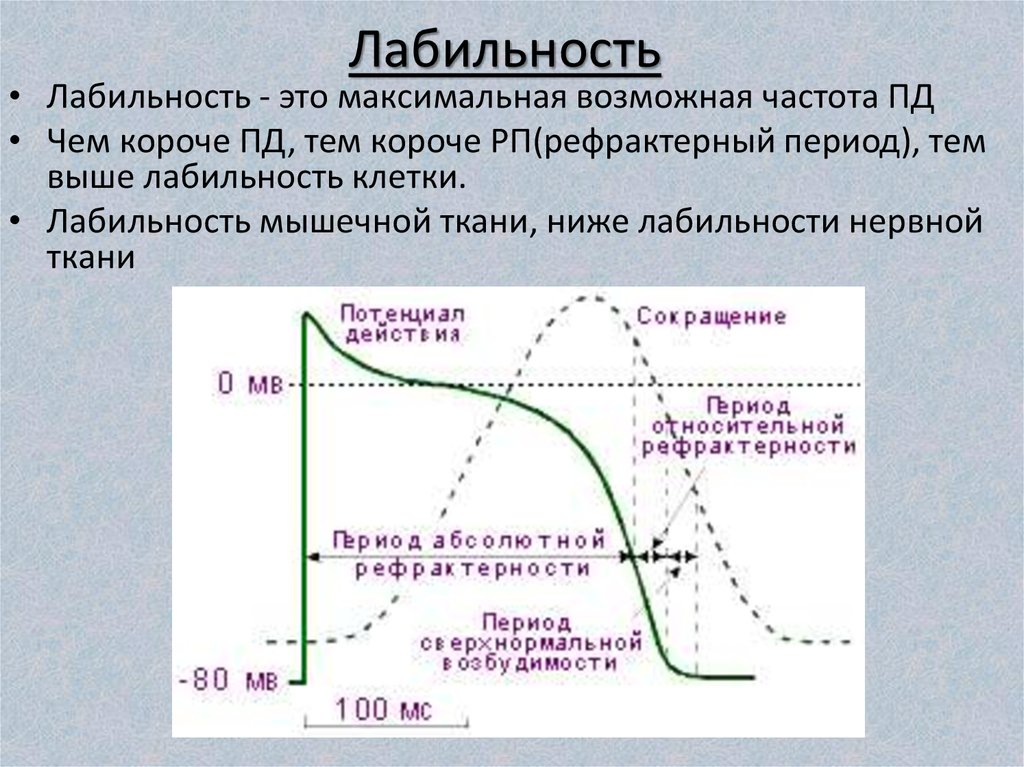
Частота импульсов возбуждения может изменяться в широких пределах. Она зависит от силы и частоты приложенного раздражения, свойств и состояния ткани.
В каждой ткани одиночный импульс возбуждения продолжается определенное время. Поэтому Н. Е. Введенский определял лабильность как скорость, с которой в ткани возникает и успевает закончиться полный период отдельного импульса возбуждения. Длительность этого возбуждения, измеряемая по продолжительности фазы абсолютной рефракторности, характеризует скорость биохимических и физиологических процессов, происходящих в ткани. Поэтому чем быстрее процессы, составляющие отдельный импульс возбуждения, тем выше лабильность и, следовательно, больше отдельных импульсов возбуждения может образоваться в ткани за определенный отрезок времени. Для измерения лабильности был предложен показатель — мера лабильности.
Мера лабильности - это максимальное число импульсов возбуждения, которые возникают за 1 с в ответ на такое же максимальное число раздражений. Высокой лабильностью обладают мякотные соматические нервы, меньшей - безмякотные вегетативные нервы. Максимальный ритм возбуждений для мякотного нерва составляет 500, для вегетативного - 200 импульсов в 1 с. Лабильность скелетных мышц выше, чем гладких. Так, максимальный ритм возбуждения для скелетных мышц - 200 импульсов в 1 с, а для гладких - в десятки раз меньше. В процессе роста и развития организма лабильность увеличивается, при старении уменьшается.
При ритмическом раздражении определенной частоты ткань не сразу отвечает соответствующим количеством импульсов возбуждения. Это связано с тем, что лабильность до начала раздражения отличается от лабильности во время раздражения — она меняется в связи с деятельностью ткани. Действие раздражителя и возникающие импульсы возбуждения меняют скорость процессов обмена веществ в ткани, которая начинает воспроизводить более частый, ранее недоступный ей ритм.
Изменение лабильности в сторону повышения или понижения по сравнению с исходным уровнем в связи с деятельностью ткани называют усвоением ритма. Лучше усваивается частый ритм при невысокой исходной лабильности. Поэтому мышечная ткань, имеющая невысокую лабильность, обладает большей способностью к усвоению ритма, чем нервная.
Оптимум и пессимум ритма и силы раздражения. Н. Е. Введенский установил, что наивысшее сокращение мышцы происходит при нанесении на нерв нервно-мышечного препарата раздражений в более редком ритме. Такой наиболее выгодный в рабочем отношении ритм раздражений был назван оптимальным (от латинского слова optimus - наилучший). При этом ритме раздражений каждый новый импульс возбуждения возникает во время фазы экзальтации, созданной предшествующим импульсом, в результате чего максимально сокращается мышца. Оптимальный ритм возбуждений меньше максимального в несколько раз. Например, максимальный ритм для двигательного нерва лягушки составляет 500, а оптимальный – 100-150 импульсов, для икроножной мышцы соответственно 120-100 и 30-50 импульсов в 1 с.
При очень частых
раздражениях, превышающих оптимальный ритм, сокращения мышцы уменьшаются и даже
совсем прекращаются - отмечают пессимум ритма раздражения (от
латинского слова pesimus - наихудший). Пессимум
возникает в том случае, когда частота раздражений превышает меру лабильности.
Под влиянием первого раздражения ткань возбуждается, затем, когда импульс
возбуждения еще не закончился, наносится повторное раздражение; оно действует
во время фазы абсолютной рефракторности. Хотя это раздражение не в состоянии
вызвать возбуждение, оно все же не остается без последствий: углубляется фаза
абсолютной рефракторности и затрудняется действие следующих за ним
раздражений. Поэтому каждое новое раздражение, посылаемое в очень частом
ритме, снижает лабильность ткани и вызывает не возбуждение, а качественно новое
состояние - торможение.

По правилу оптимума и пессимума ритма раздражений мышца сокращается и при действии раздражителя различной силы. Если увеличивать силу тока, сохраняя неизменной частоту раздражений, то происходит постепенное возрастание тетануса до максимальной высоты - оптимум силы, после чего он начинает снижаться и даже совсем исчезает - пессимум силы, когда сила тока будет чрезмерной. Это объясняется тем, что с увеличением силы раздражения импульсы возбуждения учащаются по сравнению с исходным ритмом вначале до оптимального, а затем до пессимального.
Эти положения, разработанные Н. Е. Введенским, получили дальнейшее развитие и уточнение в результате раскрытия механизма синаптической передачи. В нервно-мышечном препарате наименьшая возбудимость и лабильность выявлены у синапса. Поэтому синапс и лимитирует частоту раздражений, передаваемых с нерва на мышцу.
Методом отведения потенциалов от мышцы в области нервно-мышечного синапса установлено следующее. Редкие или оптимальные по частоте импульсы проводятся синапсом без изменений. При этом каждый импульс попадает в фазу убывания постсинаптического потенциала, вызванного предыдущим импульсом. При частом раздражении постсинаптические потенциалы суммируются, что приводит к стойкой деполяризации постсинаптической мембраны и развитию блока проведения - возникновению пессимума. Это явление часто называют пессимальным торможением или торможением Введенского.

Если инактивировать определенным химическим веществом (синап-тическим ядом) холинэстеразу, разрушающую медиатор ацетилхолин, то в области синапса будет накапливаться ацетилхолин, выделяемый нервным окончанием при каждом импульсе, и возникнет пессимум. При частых импульсах проведение их может быть блокировано еще на пути к синапсу в тонких пресинаптических окончаниях, обладающих более низкой лабильностью, чем нервное волокно.
Явления оптимума и пессимума - общебиологические свойства живой ткани, образующиеся в ответ на любые физические или химические воздействия. Умеренные раздражения оказывают оптимальное (возбуждающее) действие, а раздражения большей силы, частоты и длительности вызывают пессимальное (тормозящее) действие на реакции различных органов как при непосредственном, так и при рефлекторном воздействии на них. Явления оптимума и пессимума отмечают не только в двигательных реакциях, но и в реакциях секреторных органов и сердечно-сосудистой системы, а также в условнорефлекторных реакциях различной сложности.
4. ФИЗИОЛОГИЯ МЫШЦ
У животных существует три вида мышц: скелетные поперечнополосатые, сердечная поперечно-полосатая и гладкие, которые различны по строению и физиологическим свойствам.
СКЕЛЕТНЫЕ ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЦЫ

Скелетные мышцы вместе со скелетом составляют опорно-двигательную систему организма, которая обеспечивает поддержание позы животного и перемещение отдельных частей тела и всего тела в пространстве. Наряду с этим скелетные мышцы и скелет выполняют защитную функцию, предохраняя внутренние органы от повреждений.
Строение. Скелетные мышцы состоят из мышечных волокон, которые объединяются в мышечные пучки. Мышечное волокно - это специализированная клетка, цилиндрической формы, длиной до 10-12 см и диаметром 10-100 микрон (мкм). Каждое мышечное волокно имеет оболочку (сарколемму) и цитоплазму (саркоплазму). В саркоплазме сосредоточены все компоненты животной клетки.

Вдоль оси мышечного волокна расположены тонкие нити — миофибриллы, а в них - протофибриллы, нити белков миозина и актина. Они являются сократительным аппаратом мышечного волокна. Миофибриллы объединяются в группы - колонки, по 4-20 штук в каждой. Каждая миофибрилла разделена Z-мембранами приблизительно на 20000 саркомеров, длиной около 2,5 мкм. На обоих концах саркомеров к Z-мембране прикреплены около 2000 тонких актиновых нитей, которые своими концами входят между миозиновыми толстыми нитями.



Механизм мышечного сокращения связан с взаимодействием актина и миозина. Взаимодействие актина и миозина тормозит система мышечных белков. На поверхности актиновых нитей имеется два белка - тропонин и тропомиозин. Поступление импульса к мышце сопровождается выходом из саркоплазматического ретикулума мышечного волокна ионов Са2 . Они взаимодействуют с белком тропонином, образуя комплекс, который толкает тропомиозин в желобки между двумя цепями актина. За счет гребковых движений головок (специального белка) миозиновых нитей актиновые нити подтягиваются на миозиновые и мышца укорачивается. Кальциевый насос транспортирует Са2+ в систему саркоплазматического ретикулума, происходит отсоединение поперечных мостиков миозина от актина, и мышца расслабляется.
Свойства. Поперечнополосатым скелетным мышцам присущи следующие основные свойства возбудимых тканей - возбудимость и проводимость, а также в определенной степени упругость, растяжимость, эластичность, пластичность. Возбуждение в белых волокнах распространяется со скоростью 12- 15 м/с, а в красных- 3-4м/с.
Возбуждение мышцы внешне проявляется в сокращении. В ответ на одиночное раздражение мышца отвечает одиночным сокращением. Оно осуществляется очень быстро (за 0,09-0,1 с). В одиночном сокращении различают три фазы: скрытую, укорочения и расслабления.
В естественных условиях к мышце поступает, как правило, не один импульс, а серия импульсов. На серию импульсов мышца отвечает длительным сокращением. Оно называется тетаническим, или длительным, сокращением. Различают гладкий тетанус, который возникает при частых ритмах раздражения, и зубчатый тетанус, возникающий при редких ритмах раздражения.
Сокращение мышцы при постоянной нагрузке, сопровождающееся одним и тем же напряжением, называется изотоническим. Сокращение мышцы, когда она развивает силу, но не может укорачиваться из-за чрезмерной нагрузки, называется изометрическим.
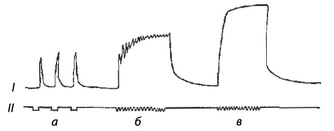
Рис. Сокращение скелетной мышцы при разной частоте раздражения: I —сокращение мышцы; II —отметка частоты раздражения; а —одиночные сокращения, б — зубчатый тетанус, в — гладкий тетанус
Поперечнополосатые скелетные мышцы относятся к произвольным мышцам, т. е. они сокращаются по воле животного. В эту группу мышц входят скелетные мышцы головы, шеи, туловища и конечностей, мышцы гортани и языка. Сократительная деятельность различных групп мышц очень разнообразна. Их согласованная деятельность обуславливает движение тела, а всякое движение вызывается сокращением большого количества мышц. Основная деятельность скелетных мышц связана с обеспечением перемещения отдельных частей и организма в целом, принятия позы. Отдельные группы мышц осуществляют свою сократительную деятельность в связи с определенными приспособительными реакциями.
Жевательные мышцы обеспечивают движение челюстей, мимические мышцы - мимику, круговые мышцы - движение губ. Затылочные, спинные, грудные, брюшные, диафрагма, межреберные мышцы участвуют в поддержании естественного положения туловища, сгибании и разгибании, повороте вправо и влево, движении тела, дыхательных движениях. Двуглавая и трехглавая мышцы передних конечностей, двуглавая, четырехглавая, икроножная мышцы задних конечностей, мышцы стопы обеспечивают сгибание и разгибание конечностей, их движение.
Работа мышц. Под работой мышц понимают удержание или перемещение тяжести за счет их сокращения. Если мышцы обеспечивают позу - то это статическая работа, если движение - это динамическая работа. Обе работы дополняют друг друга.
Сокращаясь, мышца действует на кость как на рычаг и производит механическую работу. Величину механической работы определяют как произведение массы груза на расстояние, на которое перемещен груз, и измеряют в кг*м.
Величина работы мышцы зависит от массы нагрузки и ритма работы. Увеличение массы перемещаемого груза повышает производимую работу, но до известного предела, после которого величина работы уменьшается. Наиболее производительной оказывается работа, совершаемая при средней нагрузке и среднем ритме сокращений. При частых или редких сокращениях, а также при слишком большой или малой нагрузке механическая работа мышц снижается. Законы средней нагрузки и среднего ритма мышц были установлены И. М. Сеченовым.
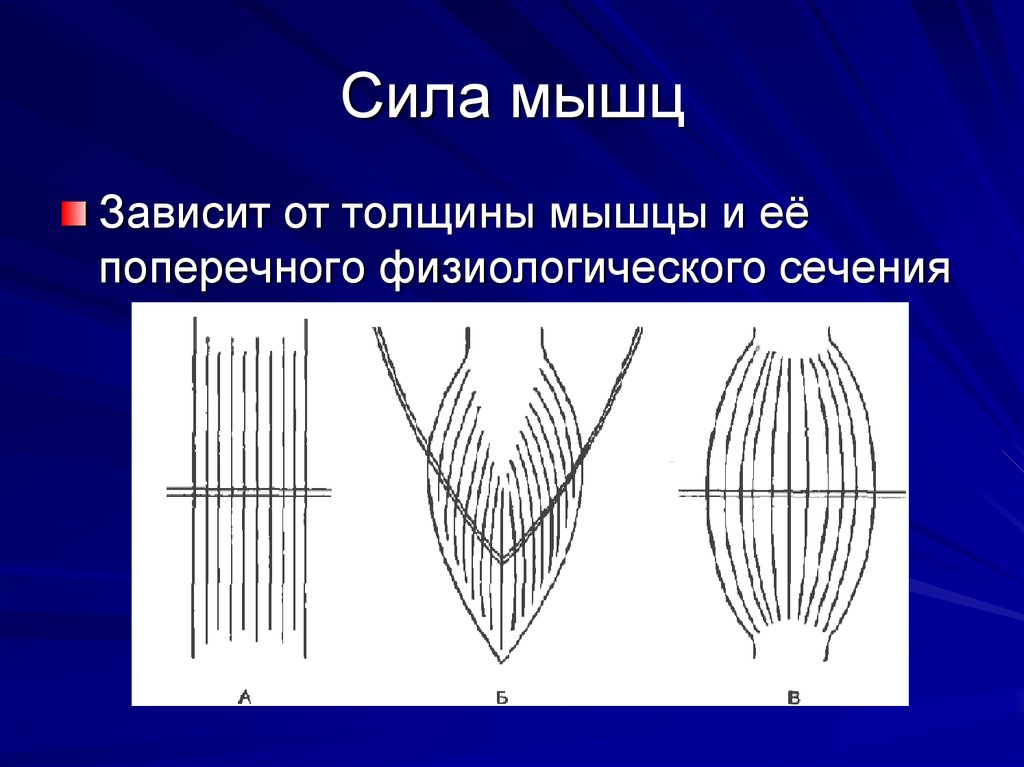
Сила мышц. Мышца характеризуется определенной силой. Она измеряется по максимальному грузу, который мышца в состоянии поднять, либо по максимальному напряжению, которое она может развить в условиях изометрического сокращения. Сила мышцы зависит от ее поперечного сечения. Отношение максимальной силы к ее анатомическому поперечнику называется относительной силой. Отношение максимальной силы мышцы к ее физиологическому поперечнику (поперечный разрез мышцы перпендикулярен ходу ее волокна) называется абсолютной силой мышцы. Мощность силы равна произведению мышечной силы на скорость укорочения.
Утомление мышц. Мышцы не могут работать беспрерывно. Длительная работа приводит к снижению работоспособности их.
Временное понижение работоспособности мышцы, наступающее при длительной работе и исчезающее после отдыха, называется утомлением мышцы. Принято различать два вида утомления мышц: ложное и истинное. При ложном утомлении утомляется не мышца, а особый механизм передачи импульсов с нерва на мышцу, называемый синапс. В синапсе истощаются резервы медиатора. При истинном утомлении в мышце происходят следующие процессы: накопление недоокисленных продуктов распада питательных веществ вследствие недостаточного поступления кислорода, истощение запасов источников энергии, необходимой для мышечного сокращения.
Утомление проявляется уменьшением силы сокращения мышцы и степени расслабления мышцы.
Если мышца на некоторое время прекращает работу и находится в состоянии покоя, то восстанавливается работа синапса, а с кровью удаляются продукты обмена и доставляются питательные вещества. Таким образом мышца вновь приобретает способность сокращаться и производить работу.
ГЛАДКИЕ МЫШЦЫ
Гладкие мышцы в организме находятся в полых внутренних органах (желудок, кишечник и др.), в кровеносных сосудах, в коже.
Строение. Основными структурно-физиологическими единицами гладких мышц являются миоциты - мышечные клетки. Их длина около 60-400 мкм, диаметр – 4-10 мкм. Клетки соединены межклеточными контактами (деомосомами). Миоцит имеет все компоненты животной клетки, ядро одно. Его сократительный аппарат, представлен нерегулярно распределенными миозиновыми и актиновыми протофибриллами. Миоциты в гладких мышцах, соединяясь между собой наружными слоями мембран в отдельных участках (нексусах), образуют «функциональный синцитий». Гладкие мышечные волокна не имеют поперечной исчерченности. Клетки укорачиваются в результате относительного скольжения нитей. Скорость скольжения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. Благодаря этому гладкие мышцы хорошо приспособлены для длительного стойкого сокращения без утомления, с меньшей затратой энергии.

Свойства. Гладким мышцам присущи те же основные свойства, что и поперечно-полосатым скелетным мышцам, но и некоторые особые свойства: автоматия, т. е. способность сокращаться и расслабляться без внешних раздражений, а за счет возбуждений, возникающих в них самих (в пейсмекерных клетках); высокая чувствительность к химическим раздражителям; выраженная пластичность (свойство сохранять приданное растяжением состояние); сокращаются в ответ на быстрое растяжение.
Гладкие мышцы относятся к непроизвольным мышцам, т. е. их сокращение не зависит от воли животного. Особенности двигательной деятельности желудка, кишечника, кровеносных сосудов и кожи в известной степени определяют физиологические особенности гладких мышц этих органов.
5. ФИЗИОЛОГИЯ НЕРВОВ
Нерв - это сложное образование, состоящее из большого числа нервных волокон, заключенных в общую соединительнотканную оболочку. Нервное волокно представляет собой отросток нервной клетки (нейрона) - дендрит или аксон.

Строение. Нервное волокно состоит из осевого цилиндра и покрывающих его миелиновой оболочки, прерывающейся через каждые 1-2,5мм (перехваты Ранвье) и Шванновской оболочки, образованными Шванновскими клетками (миелоцитами). Нервные волокна, не имеющие миелиновой оболочки, называют безмякотными, а имеющие - мякотными. Осевой цилиндр содержит аксоплазму, пронизанную нейрофибриллами и микротрубочками, покрыт плазматической мембраной. Каждая структура нервного волокна несет определенную физиологическую роль: осевой цилиндр проводит возбуждение; миелиновая оболочка обеспечивает изолированное проведение возбуждения от других нервных волокон и участвует в питании осевого цилиндра; нейрофибриллы и микротрубочки обеспечивают ток аксоплазмы. Все нервные волокна по строению подразделяют на миелиновые и безмиелиновые, а по физиологическому значению (структурным особенностям и скорости проведения возбуждения) - на тип A (Aα, Аβ, Aγ, Aδ), толстые миелиновые, быстропроводящие афферентные и эфферентные, тип В, вегетативные преганглионарные мякотные и тип С, безмякотные, тонкие, медленнопроводящие постганглионарные вегетативные нервные волокна и афферентные волокна от рецепторов тепла, давления, боли.
Свойства. Основные свойства нерва, нервного волокна: возбудимость и проводимость. Возбуждение в виде потенциалов действия проводится от рецепторов в центральную нервную систему по афферентным волокнам, от ЦНС к периферическим исполнительным органам по эфферентным нервным волокнам. Возбуждение проводится по нервному волокну в случае анатомической и физиологической целостности, изолированно по каждому волокну.
Нерв относительно неутомляем, т. к. тратит мало энергии (в основном на работу Na/K насоса).
6. НЕРВНО-МЫШЕЧНАЯ ПЕРЕДАЧА ВОЗБУЖДЕНИЯ. СИНАПС
Передача возбуждения с нервного волокна на нервную, мышечную и железистую клетки осуществляется через специальное структурное образование, с помощью особого механизма и происходит в результате выделения нервными окончаниями химических соединений - медиаторов (передатчиков) нервного импульса. Роль медиатора у животных в скелетных мышцах играет ацетилхолин.
Структурно-физиологическое образование, обеспечивающее передачу возбуждения с нервного волокна на иннервируемую им клетку (мышечную, нервную или железистую), называется синапсом. В зависимости от расположения и роли различают синапсы: аксоаксональные, аксодендрические, аксосоматические, возбуждающие, тормозные, смешанные - с электрической и химической передачей.
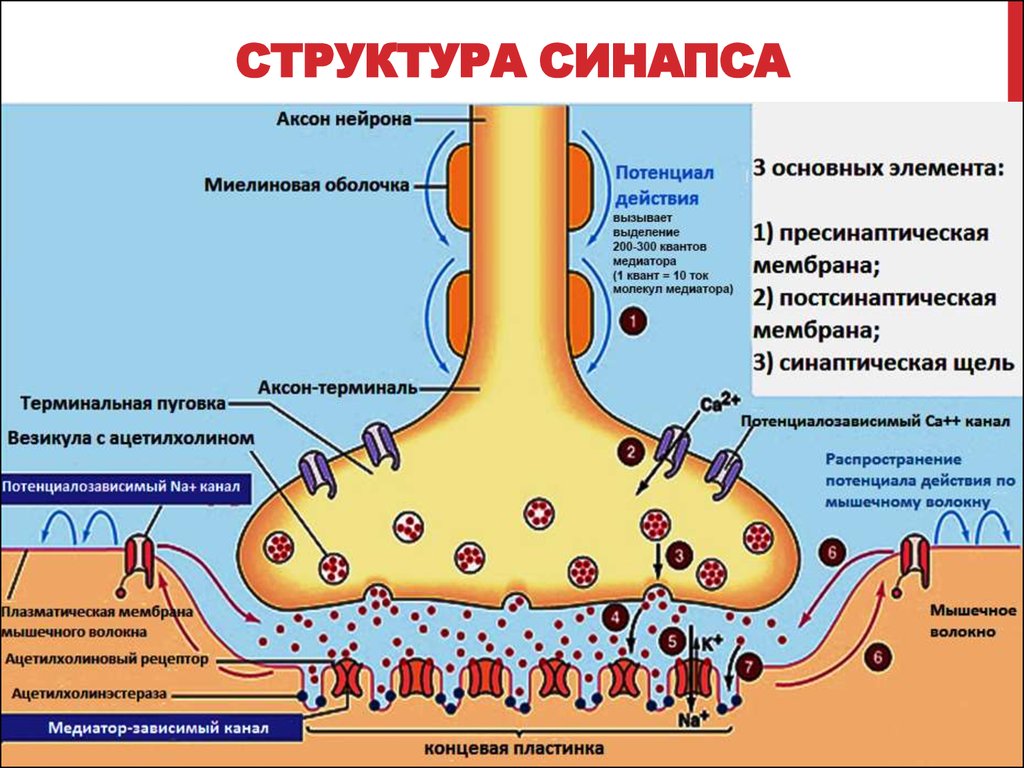
Синапсы. Состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели.
Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части расположены пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в «пузырьках» (гранулах).
Постсинаптическая мембрана - специальное образование, утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Имеет электрически возбудимые ионные каналы и поэтому способна к генерации потенциала действия; специальные белковые структуры - рецепторы, воспринимающие действие специфических химических веществ - медиаторов.
Синаптическая щель - пространство между пре- и постсинаптическими мембранами, которое заполнено жидкостью, близкой по составу к плазме крови; ее размер колеблется от 50 до 500 нанометров (нм).
Механизм передачи возбуждения через синапс. Возбуждение распространяется по нервному волокну в виде потенциала действия (нервного импульса) и, достигнув пресинаптической мембраны, вызывает ее деполяризацию, что приводит к открытию кальциевых каналов. Ионы Са2+ входят внутрь нервного окончания и способствуют освобождению медиатора из «пузырьков» и выходу его в синаптическую щель. Медиатор быстро диффундирует через щель и воздействует на постсинаптическую мембрану - взаимодействует с рецептором (для ацетилхолина - с холинорецептором, для норадреналина - с адренорецептором и др.). На взаимодействие медиатора с рецептором мембрана отвечает изменением проницаемости для ионов Na+ и К+, что приводит к ее деполяризации, возникновению потенциала действия и генерации возбудительного постсинаптического потенциала (ВПСП). Под влиянием этого потенциала происходит деполяризация соседних с синапсом участков мембраны. Потенциал действия распространяется по всему органу.
Медиаторы поступают в синаптическую щель не только при возбуждении, но и в покое. В покое выделяются малые порции («кванты») медиатора, а под влиянием нервного импульса - одновременно выбрасываются значительные количества таких «квантов». Избыток медиатора разрушают специальные ферменты: холинэстераза - ацетилхолин, моноаминооксидаза и катехол-О-метилтрансфераза - норадреналин и др.
Свойства синапсов. Синапсы обладают рядом следующих специфических свойств:
Одностороннее проведение возбуждения - возбуждение проводится через синапс только в одну сторону (в одном направлении) - с нерва (аксона) на мышцу (другой орган).
Замедление проведения возбуждения - возбуждение через синапсы проводится с задержкой в сравнении с нервным волокном. Это объясняется тем, что проведение возбуждения - многоэтапный процесс, т. е. время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, активацию мембраны, рост потенциала действия до пороговой величины.
Утомление - связано с уменьшением резерва медиатора при длительном поступлении импульсов.
Низкая лабильность - обусловлена тем, что проведение возбуждения через синапс сопряжено с затратой относительно большого количества времени.
В возбуждающих синапсах медиаторами являются норадреналин, донамин, сербонин. Ацетилхолин – универсальный медиатор.
В гладких мышцах и в центральной нервной системе имеются не только возбуждающие, но и тормозные синапсы. Нервные импульсы высвобождают из нервных окончаний в них тормозной медиатор (в кишечнике, бронхах - норадреналин, в мышечных клетках водителя ритма сердца - ацетилхолин, в центральной нервной системе - гамма-аминомасляная кислота и др.).
Отличительные особенности тормозного синапса:
1. Синаптическая щель тормозного синапса уже и составляет 20 нм, у возбуждающего – 30 нм;
2. Постсинаптическая мембрана тормозного синапса более толстая и плотная;
3. Пресинаптическая мембрана тормозного синапса содержит меньше пузырьков медиатора;
4. Медиаторами в тормозных синапсах являются глицин, гаммааминомасляная кислота (ГАМК), иногда – ацетилхолин.
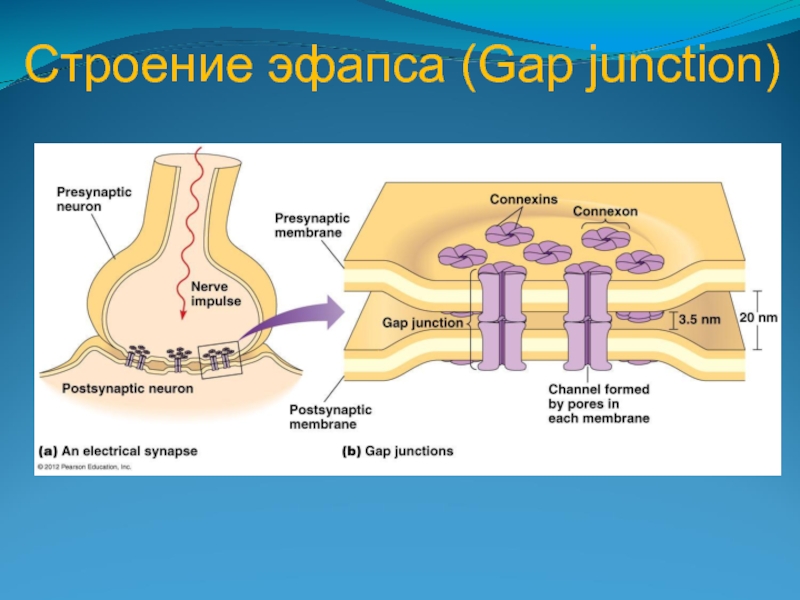
Относительно редко встречаются электрические синапсы (эфапсы); потенциал действия передается без химического посредника, в обоих направлениях и практически без задержки. Передача возбуждения в этих синапсах называется эфаптической.